 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(5) July 2001, pp. 627-634 Spatio-temporal Dynamics and Transition from Asymptotic Equilibrium to Bounded Oscillations in Chrysomya albiceps (Diptera, Calliphoridae) Wesley Augusto Conde Godoy/+, Fernando José Von Zuben*,Cláudio José Von Zuben**, Sérgio Furtado dos Reis*** Departamento de Parasitologia, IB,
Universidade Estadual Paulista, Rubião Junior, 18618-000 Botucatu, São Paulo,
Brasil *Departamento de Engenharia de Computação e Automação Industrial ***Departamento
de Parasitologia, IB,Universidade Estadual de Campinas, Campinas, SP, Brasil
**Departamento de Zoologia, IB, Universidade Estadual Paulista, Rio Claro, SP,
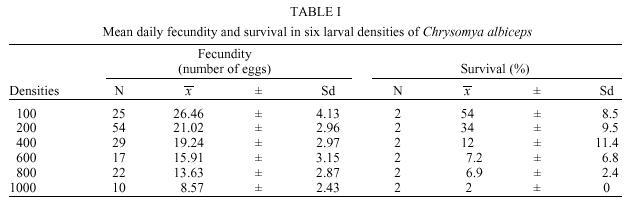
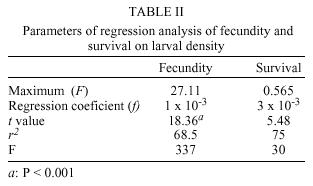
Brasil Received 15 August 2000 Code Number: oc01095 The sensitivity of parameters that govern the stability of population size in Chrysomya albiceps and describe its spatial dynamics was evaluated in this study. The dynamics was modeled using a density-dependent model of population growth. Our simulations show that variation in fecundity and mainly in survival has marked effect on the dynamics and indicates the possibility of transitions from one-point equilibrium to bounded oscillations. C. albiceps exhibits a two-point limit cycle, but the introduction of diffusive dispersal induces an evident qualitative shift from two-point limit cycle to a one fixed-point dynamics. Population dynamics of C. albiceps is here compared to dynamics of Cochliomyia macellaria, C. megacephala and C. putoria. Key words: spatial dynamics - sensitivity analysis - Chrysomya albiceps The invasion of new habitats by organisms is an important ecological phenomenon since invading species generally have tremendous ecological and economic impact on new areas (Hengeveld 1989, Andow et al. 1993, Kareiva 1996). Biological invasions can take place in different ways including invasions into patchy environments and by stratified diffusion both in short and long-range dispersal (Shigesada & Kawasaki 1997). The consequences of an invasion may vary from competition for food or space between invading and native species, to invasion of parasites and the spread of epidemic diseases (Hengeveld 1989, Shigesada & Kawasaki 1997). Invader populations depend on physical and biological factors for success in invasion and colonization process (Stiling 1996). Among the main biological factors associated to population growth as well as success in colonization, persistence and extinction of populations in new areas deserve special attention since the future of the invading species in its new habitat depends basically on the period of time that they stay in the new habitat (Hengeveld 1989, Caughley & Gunn 1996, Hanski 1998). The temporal change in the population density of organisms at certain spatio-temporal coordinates can occur as a result of births, deaths, immigration and emigration (Rhodes & Odum 1996, Hastings 1997). Methodologically it is simpler to study demographic processes only in a temporal context because they are essentially an one-dimensional problem (Turchin 1998). Migration, however, involves two scales - temporal and spatial - rendering its study intrinsically more complex (Schneider 1989). Ecologists have realized that movement is a critical but little understood process affecting population numbers (Ranta et al. 1998). Movement can subtract or add individuals to a population, can alter the outcome of species interactions, provide crucial genetic variability and rescue population from extinction (Kendall & Fox 1998, Turchin 1998). However, to know whether any of these postulated effects are of practical importance, we need to be able to investigate dispersal. Around twenty years ago, three species of blowflies originary from Africa and Asia, Chrysomya albiceps, C. megacephala and C. putoria have become established in Brazil (Guimarães et al. 1978, 1979, Baumgartner & Greenberg 1984). This invasion has culminated with the sudden decline in population numbers of an ecologically similar species, Cochliomyia macellaria (F.), which is native to the Americas (Greenberg & Szyska 1984). Some life-history aspects of the introduced species can be implied in the displacement of the native species C. macellaria, since C. albiceps is a species that exhibits a predatory behavior on larvae of other species during the larval stage (Coe 1978, Gagné, 1981, Erzinçlioglu & Whitcombe 1983, Faria et al. 1999). This behavior has been studied in details by a choice and no-choice experiment, where there are strong evidences to believe that C. albiceps larvae prefer C. macellaria larvae instead of C. megacephala and C. putoria (Faria et al. 1999). In addition Chrysomya species have shown higher competitive ability than C. macellaria in experimental and natural conditions (Wells & Greenberg 1992a,b,c, Reis et al. 1999). Besides the biological invasion problem, these species are important mechanic vectors of diseases and causers of facultative myiasis in humans and animals (Zumpt 1965, Greenberg 1971, 1973, Baumgartner & Greenberg 1984, Wells 1991). We have initiated a research program as an attempt to better understand the blowflies invasion process (Godoy et. al. 1996, 1997, Reis et al. 1996). In this program we have integrated mathematical and biological approaches in order to address questions involving spatio-temporal dynamics. By use of mathematical models of population growth, the dynamic behavior of two invader species, C. megacephala and C. putoria, and of the native species C. macellaria was analyzed and the results indicate that the introduced and native species differ markedly in their equilibrium dynamics (Godoy et al. 1993, 1996, 1997, Von Zuben et al. 1993, Reis et al. 1996, Teixeira et al. 1998). Application of the mathematical model using density-dependent parameters such as fecundity and survival derived from experimental populations showed that C. megacepahala and C. putoria exhibit stable oscillations with numbers fluctuating between two points in sucessive generations (Godoy et al. 1993, Von Zuben et al. 1993), whereas in C. macellaria the dynamics is characterized by damping oscillations in population size leading to one fixed point equilibrium (Reis et al. 1996, Godoy et al. 1997). In this paper we investigate the theoretical dynamics of experimental populations of C. albiceps as an attempt to understand its population biology and compare the results obtained with those observed by Reis et al. (1996), Godoy et al. (1997) and Teixeira et al. (1998). We specifically, analyzed the population dynamics of C. albiceps by means of a mathematical model that incorporates fecundity and survival as density-dependent demographic parameters in discrete time. We also investigated the sensitivity of these parameters to changes in the magnitude of its values and analyzed the dynamic behavior in a spatial structure based on the coupled lattice maps formalism, in order to investigate if the population dynamics of these species is influenced by spatial migration. MATERIALS AND METHODS Laboratory population of C. albiceps - Specimens of C. albiceps were collected in the vicinities of the campus of Universidade Estadual Paulista, Botucatu, São Paulo, Brazil. Adult flies were maintained in laboratory conditions in cages (30x30x30cm) covered with nylon at 25 ± 1°C and were fed water and sugar ad libitum. Adult females were fed fresh beef liver to permit the complete development of the gonotrophic cycle (Linhares 1988). The experiment was performed using the generation F2, which is progeny of one generation which had its life cycle completed in the laboratory. Exploitative intraspecific competition among immatures, which is known to occur under natural conditions (de Jong 1976, Lomnicki 1988) was established in the laboratory by setting up six replicated larval densities, developing in 20 g of ground beef, ranging from 100 to 1,000 larvae per vial at intervals of 200. This range of densities is suitable to simulate intraspecific competition process in C. albiceps since it produces decrease in demographic parameters as a function of density (Godoy et al. 1993, Von Zuben et al. 1993, Reis et al. 1994, 1996). Fecundity was measured counting the number of eggs per female and expressed as the average of daily egg output, which is based on the length of the gonotrophic cycle of Chrysomya species at 25°C (Avancini & Prado 1986, Linhares 1988). Maximum sample size for estimation of fecundity was 30 females per vial. Sample sizes smaller than 30 in some vials were due to either low immature survival rates or incomplete ovarian development. Survival was estimated as the number of adults emerging from each vial. Mathematical model - The mathematical model developed by Prout and McChesney (1985) was applied to investigate the dynamics of laboratory populations of C. albiceps. This model is based on a finite difference equation that models population dynamics considering the number of immatures, eggs or larvae, in succeeding generations, nt+1 and nt. Prout and McChesney's model incorporates two density-dependent processes, the variation of fecundity and survival as function of density of immatures, nt. The recursion is written in the nonlinear finite difference equation nt+1 = ½ F* S* e _ (ƒ+ s) nt nt (1) where F* and S* are the intercepts in the regression analysis of fecundity and survival as a function of larval density. These parameters describe the theoretical values for maximum fecundity and survival, respectively. The factor of ½ indicates that only half of the population are adult females which contribute with eggs to the next generation. The values of ƒ and s are regression coefficients that estimate the slope of fecundity and survival on the density of immatures. The exponential function was used because it fitted the C. albiceps data equal to or better than linear and hyperbolic functions and, the linear regression is known to produce larger slopes in absolute magnitude that produces larger eigenvalues which do not accurately describe the model dynamics at carrying capacity (Mueller 1986). In addition the decrease in fecundity as a function of density of immatures can be viewed biologically as a Poisson process, which is decribed by an exponential function (Rodriguez 1989). The effect of parameter variation on the population dynamics of C. albiceps was investigated through simulations varying maximum fecundity (F*) and survival (S*), intercepts in the regression analysis. All simulations were carried out with Matlab (Moler et al. 1987). In the simulations reported here, fecundity was arbitrarily allowed to vary up to a mean daily egg output of 40 eggs (the maximum value of fecundity observed for females of this species; pers. obs.). Spatial structure
was incorporated to equation (1) employing the physical formalism of coupled
map lattices (Bascompte & Solé 1994). The spatial dimension was incorporated
as a discrete lattice of points with the populations arranged on the nodes of
this square lattice and the population in each node is linked with dispersal
to the four nearest neighbors. The spatial structure is incorporated to equation
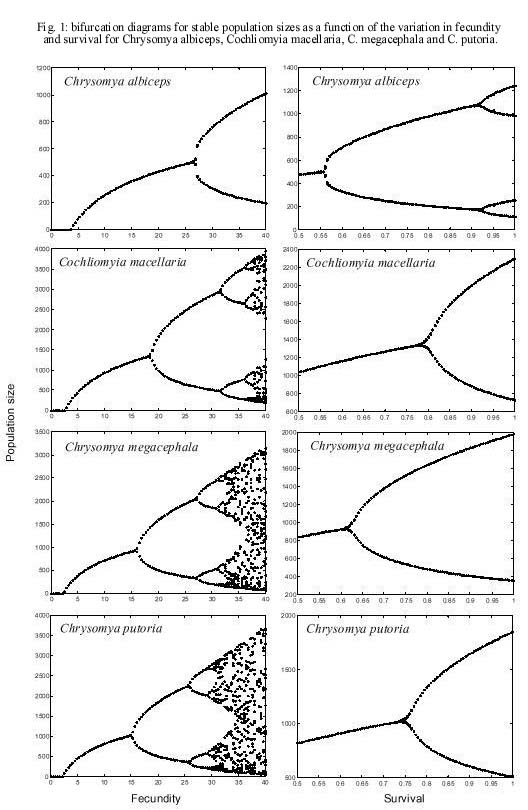
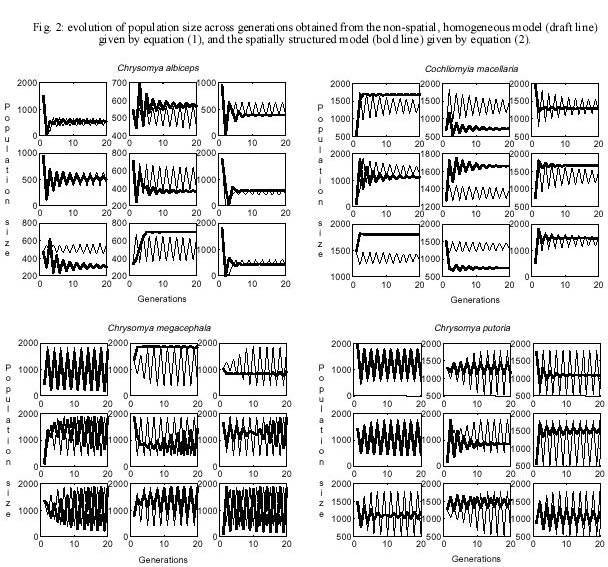
(1) yielding an equation for the spatio-temporal dynamics as The findings for the temporal and spatial dynamics of C. albiceps obtained in this paper are compared and discussed with those obtained for C. megacephala, C. putoria and C. macellaria, which are described in Godoy et al. (1996, 1997) and Reis et al. (1996) using the same mathematical formalism. RESULTS Survival and fecundity of C. albiceps decreased as function of density of immatures and this decrease was statistically significant (Tables I, II). Slopes of the exponential regression for fecundity and survival were significantly different from zero (P < 0,001) (Table II). Theoretical values for maximum fecundity (F*) and survival (S*), and the regression coefficients (Table II) describing the dependence of fecundity (f) and survival (s) upon density of immatures were incorporated to equation (4), which can be written as nt+1 = ½ [(27.11 e _ 0.001 nt )(0.56 e _ 0.003 nt) nt ] (4) The qualitative dynamics of C. albiceps can be assessed by the eigenvalue associated with equation (1), which is larger than 1 in modulus (l = -1.0398) and has so two points stable equilibrium. The dynamic behavior observed for C. albiceps was derived from the application of fixed parameter values to the non-linear difference equation (1). Bifurcation diagrams show that increasing values for fecundity produce qualitative changes in the dynamics of the C. albiceps (Fig. 1) and, for values of fecundity larger than 26 eggs, there is apparently a transition in the dynamics from stable equilibrium to two point limit cycle. When survival is analyzed, the upper limit is 1.0, which represents a theoretical maximum viability of 100%. In this case we can see that the effect of survival on the dynamics of C. albiceps is much more noticeable than that of fecundity, since two bifurcations were realized (Fig. 1). The consequences of diffusive dispersal for the population dynamics of C. albiceps are illustrated in Fig. 2. The oscillations in population numbers as a function of time measured in generations is shown, both for the non-spatial homogeneous model, equation (1), and the spatially structured model, equation (2) (Fig. 2). Our simulations suggest that the linkage between populations introduced by diffusive dispersal produces a qualitative effect on the local dynamics of C. albiceps from two point limit cycle to stable equilibrium (Fig. 2). In addition, spatial structure introduced a small quantitative effect in the population dynamics, characterized by the change in the spectrum of the oscillations. DISCUSSION The mathematical analysis performed in this study reveals that C. albiceps shows a dynamic behavior represented by a limit cycle of two points, since l = -1,0398 (larger than one in module). This behavior was also observed in C. megacephala and C. putoria (Godoy et al. 1993, Von Zuben et al. 1993), but it differs from the native species, C. macellaria, that shows damping oscillations in population size leading to one fixed point equilibrium (Reis et al. 1996). We noted, however, that the value of l obtained for C. albiceps is very close to one if compared to the other values observed for C. megacephala, l = -1.3761 and C. putoria, l = -1.2399 (Godoy et al. 1996). This result suggests that C. albiceps is a species apparently more susceptible to changes in its dynamic behavior than C. megacephala and C. putoria. Stability of equilibrium has been extensively investigated in insect populations (Dennis et al. 1995, Costantino et al. 1995, 1997, 1998, Cushing et al. 1998) and perhaps the most important question arising in this context be what governs the transitions from stable to unstable equilibria. The effects produced by the manipulation of the parameter values are well known in theoretical population models (May & Oster 1976, Edelstein-Keshet 1998) and evidences for such changes come from theoretical and empirical studies (Cavalieri & Koçak 1995, Costantino et al. 1995, 1997, Denis et al. 1995, 1997). Experiments designed to test prediction of population models have shown that shifts in dynamical behavior can occur, from stable point equilibria, to stable limit cycles, to aperiodic cycles in response to changes in the rate of adult stage mortality (Costantino et al. 1995, 1997, Denis et al. 1995, 1997). In our study the simulations revealed that the transition from asymptotic stable equilibrium to bounded oscillations occurs with an increase in fecundity and survival. It is interesting to note that the results of this study indicate a more frequent variation of dynamic behavior when the survival values are manipulated. Recently, Godoy et al. (1996) used the same technique to analyse the transition of dynamic behavior in C. megacephala, C. putoria and C. macellaria and observed that the effect of survival on the dynamics of these species is much less noticeable than of fecundity, since no more than one bifurcation was obtained in the three species studied (Fig. 1). Fecundity was shown in the simulations to be the most important demographic parameter to bring about shifts in the dynamic behavior of experimental populations of C. megacephala, C. putoria and C. macellaria (Godoy et al. 1996). The coupled map lattices suggest that the linkage between populations introduced by diffusive dispersal produces a qualitative effect on the local dynamics of C. albiceps from two point limit cycle to stable equilibrium (Fig. 2). These results confirm the findings by Godoy et al. (1997) which revealed that the linkage between populations introduced by diffusive dispersal may produce qualitative and quantitative effects on the local dynamics of C. megacephala, C. putoria and C. macellaria (Fig. 2). The dynamics system in which C. albiceps is inserted gives a wide range of interesting questions since it apparently balances between stability and periodic behavior. This apparent ambiguity as a function of both the parametric manipulation and the spatial structure introduced in the model, places C. albiceps between C. macellaria, the native species that exhibits stable dynamics and the other introduced species, C. megacephala and C. putoria, that show two point limit cycle (Reis et al. 1996). We consider this point interesting because in this sense C. albiceps is a species that shows a dynamic behavior more susceptible to changes than C. megacephala and C. putoria. We do not know if this characteristic is advantageous to C. albiceps, however it has proved to be a species with conspicuous abilities under competitive stress, what probably have influenced negatively the structure of the community where it lives (Ullyett 1950, Coe 1978, Faria et al. 1999). The spread of invading organisms is the outcome of complex interactions involving the dynamics of populations at temporal and spatial scales (Hengeveld 1989). The importance of the spatial component has always been recognized (Lewis & Kareiva 1993), although recently a stronger emphasis has been placed on spatial structure as result of theoretical developments that have established a correlation between migration and dynamic behavior of populations (Comins et al. 1992, Allen et al. 1993, Hastings 1993, Bascompte & Solé 1995, Ruxton 1995, Kendall & Fox 1998). The results found in these investigations show two opposite ways, on the one hand the complex or chaotic temporal dynamics can be induced by the presence of the spatial dimension (Comins et al. 1992, Pascual 1993) and on the other hand, simple periodic dynamics may be induced by the presence of the diffusive dispersal (Hastings 1993, Csilling et al. 1994). Our simulations indicate that the spatial dimension induces an overall stabilizing effect on the dynamics of C. albiceps. This paper, together with the previous focus the connection between population dynamics and the invasion of blowflies (Reis et al. 1996, Godoy et al. 1996, 1997), seem to confirm the findings of Costantino et al. (1995, 1997) and Allen et al. (1993), which have taken together a new conceptual thread connecting sensitivity to demographic changes, qualitative changes in dynamic behavior and extinction rates. In addition, the peculiar characteristics found in C. albiceps, which seem to have implications to population dynamics, open a series of new perspectives for studies of equilibrium dynamics. ACKNOWLEDGMENTS To the anonymous reviewer for making suggestions that improved the clarity of the manuscript. REFERENCE
CJVZ, FJVZ and SFR has been supported by research fellowships from Conselho Nacional de Desenvol-vimento Científico e Tecnológico. Copyright 2001 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc01095t2.jpg] [oc01095f2.jpg] [oc01095t1.jpg] [oc01095f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}