 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(5) July 2001, pp. 673-678 In Vivo Lymphocyte Activation and Apoptosis by Lectins of the Diocleinae Subtribe Theolis Barbosa, Sérgio Arruda, Benildo Cavada*, Thalles Barbosa Grangeiro*, Luiz Antônio Rodrigues de Freitas, Manoel Barral-Netto/+ Centro de Pesquisas Gonçalo
Moniz-Fiocruz, Rua Valdemar Falcão 121, 40295-001 Salvador, BA, Brasil
*Centro de Ciências, Universidade Federal do Ceará, Fortaleza,
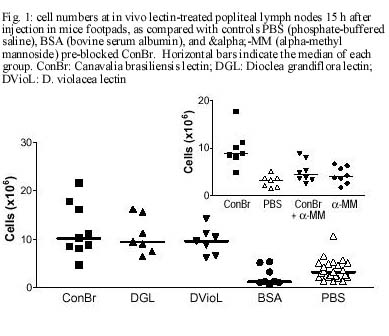
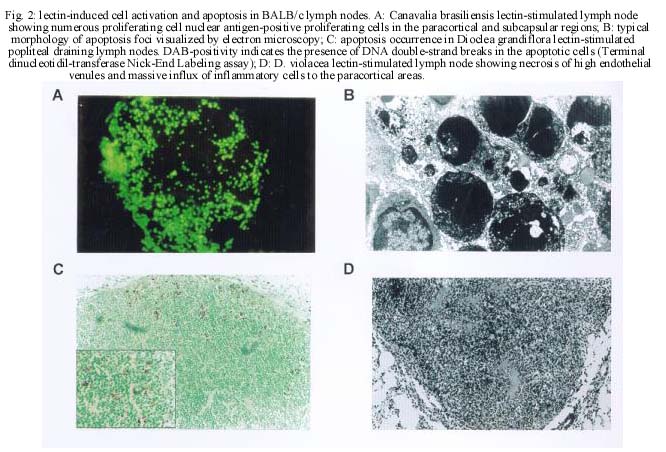
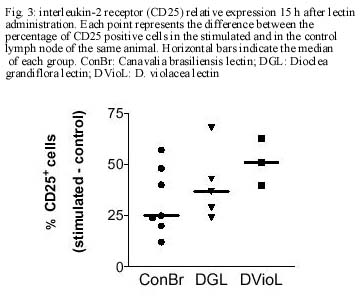
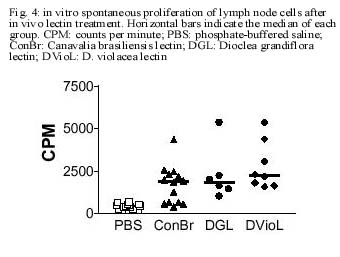
CE, Brasil Received 24 October 2000 Code Number: oc01105 This paper reports the overall effects of three lectins, extracted from Canavalia brasiliensis, Dioclea violacea, and D. grandiflora, on BALB/c mice popliteal draining lymph nodes. These lectins have presented high stimulatory capacity on lymph node T cells. Additionally, they were able to induce apoptosis and inflammation (frequently associated with high endothelial venule necrosis). The data presented here suggest that the Diocleinae lectins studied can stimulate in vivo T cell activation and apoptosis, as well as present important side effects. Key words: adjuvants - apoptosis - Diocleinae - lectin In the present work we studied the in vivo stimulatory capacity of three lectins isolated by our group (Moreira et al. 1983, 1996, Moreira & Cavada 1984, Ainouz et al. 1987), extracted from Brazilian legume species of the Diocleinae subtribe: Canavalia brasiliensis (ConBr), Dioclea grandiflora (DGL) and D. violacea (DVioL). These lectins have previously been shown to induce proliferation and interferon-gamma production by peripheral blood mononuclear cells in culture (Barral-Netto et al. 1992). Moreover, they presented a higher stimulatory capacity than concanavalin A (ConA), in spite of having over 80% homology with this lectin (Moreira et al. 1983, Barcellos et al. 1993, Cavada et al. 1993). We have evaluated their effects on BALB/c mice popliteal draining lymph nodes after subcutaneous injection of footpads. Lectin stimulation was found to be associated to apoptosis induction. Other effects observed included inflammation and high endothelial venule necrosis. MATERIALS AND METHODS Lectin-treatment of mice - The methods for lectin purification were published elsewhere (Moreira et al. 1983, 1996, Moreira & Cavada 1984). Lectin preparations were tested to exclude LPS contamination (data not shown). Inbred BALB/c mice of both sexes, 8-14 weeks of age, were used in all experiments. Animals were injected s.c. with 50 µg of lectin or bovine serum albumin (BSA, Sigma, St. Louis, MO) in 20 µl of phosphate-buffered saline in the left hind footpad. As controls, the right hind footpads were injected with 20 µl of PBS. As a lectin inhibition control, a ConBr solution with the lectin concentration mentioned above was prepared in phosphate-buffered alpha-methyl mannoside (a-MM, Sigma) 1M, left for 1h at room temperature and administered in the left footpad of eight animals, with a-MM 1 M in the right footpads for comparison. Animals were sacrificed 15 h after lectin injection. In vitro proliferation assays - Single cell suspensions in PBS were obtained from the popliteal draining lymph nodes of the mice and counted in a hemocytometer. For spontaneous proliferation assessment, lymph node cells were cultivated in RPMI medium (GibcoBRL, Rockville, MD) supplemented with 10% fetal calf serum (Sigma), penicillin, streptomycin (100 U/ml and 100 mg/ml, respectively; GibcoBRL), and 2-mercapto-ethanol (50 µM; Sigma) for 24 h in 96-well flat bottom plates (Corning, NY) in humidified atmosphere (37°C, 5% CO2). [H3]-thymidine (Amersham, UK) was added at 1 µCi per well and [H3]-thymidine incorporation was evaluated in triplicate by liquid scintillation radioactivity counting (b-Counter1409 DSA; Wallac, Gaithersburg, MD). IL-2 receptor (IL-2R) expression evaluation - One million lymph node cells were stained with anti-CD25 (IL-2R) FITC conjugated monoclonal antibody (AMT-13 clone, Sigma) in PBS with 1% bovine serum albumin (BSA) and 0.1% sodium azide (Sigma). Controls included isotype control stained (Pharmingen, San Diego, CA) and unstained samples evaluated in triplicate. Flow cytometry analyses were carried out in a FACSort flow cytometer (Becton-Dickinson Immuno-cytometry Systems, Franklin Lakes, NJ). Morphological evaluation of treated lymph nodes - Popliteal draining lymph nodes from lectin injected mice were fixed in 10% buffered formaldehyde (Sigma) for at least 12 h, embedded in paraffin and stained with hematoxylin and eosin (Sigma). Electron microscopy - Small fragments from the draining lymph nodes were immediately fixed in 2% glutaraldehyde (Sigma) in 0.1 M cacodylate buffer (Sigma) for 1-2 h, postfixed in osmium tetroxide (Sigma), washed in buffer, dehydrated in graded acetone and embedded in epon resin. Semi-thin sections were cut on a Reichert-Jung Ultracut E microtome with glass knives, mounted on glass slides and stained with methylene blue- azurII to identify areas of lymphocyte apoptosis. Ultra-thin sections were cut with a diamond knife, mounted on copper grids and contrasted with uranyl acetate and lead citrate. Observations were carried out on a Zeiss (Germany) EM-109 electron microscope at 50 kV. In situ evaluation of PCNA expression - The PCNA (Proliferating Cell Nuclear Antigen)-positive proliferating cells were visualized by immunofluorescence in 3-5 mm frozen lymph node sections permeabilized with saponin (Sigma) 0.1% in PBS, using anti-PCNA FITC-conjugated antibody (clone 19F4, Coulter, Fullerton, CA). The sections were photographed at the epifluorescence microscope (Axioskop; Zeiss, Germany). In situ detection of apoptotic cells through the TUNEL assay - Apoptosis occurrence in the lymph nodes was demonstrated in 3-5 mm thick paraffin sections through the TUNEL (Terminal di-nucleotidil-transferase Nick-End Labeling) technique, using the Apoptag Plus kit (Oncor, Gaithersburg, MD). Statistical analyses - Data obtained from lymph node cell counts were compared using parametric one-way ANOVA followed by Student-Newman-Keuls Multiple Comparison Test and Student's t test. Proliferation assessment assays and flow cytometric IL-2R expression evaluations were compared through Kruskal-Wallis one-way ANOVA followed by Dunn's post test. We have used the GraphPad Prism software (v. 2.00, GraphPad Software Inc., San Diego, CA). RESULTS Lectins (50 µg/footpad) promoted a remarkable enlargement of BALB/c mice popliteal lymph nodes 15 h after the subcutaneous injection into the footpads of mice. This augmentation was due to the expansion of paracortical and interfollicular regions (data not shown), and was accompanied by an increase in lymph node total cell counts, as expressed in Fig. 1. The number of cells in stimulated lymph nodes was significantly higher than in saline-injected controls, but no significant differences were found among the lectins studied (p=0.3430). Administration of bovine serum albumin (BSA), an unrelated protein, to murine footpads did not produce a significant increase in lymph node cell counts, as compared to saline injection (p=0.2896), and such counts were significantly lower then those obtained for lectin-stimulated nodes (p<0.002). Pre-blockage of ConBr with α-MM has significantly inhibited lectin stimulatory activity (p<0.01, Fig. 1 insert). Pre-blocked ConBr stimulated lymph nodes were not significantly different from α-MM-treated ones or PBS controls (p>0.05). Upon lectin stimulation T cell areas (parafol-licular, interfollicular and subcapular regions) were predominantly expanded, exhibiting abundant lymphoblasts and dividing cells. In contrast, the follicles maintained the unstimulated appearance, with small cells and lack of germinative centers (data not shown). The in situ evaluation of PCNA (proliferating cell nuclear antigen) expression reveals abundance of cells undergoing proliferation in the stimulated lymph nodes, especially in T cell areas (Fig. 2A). In contrast, PBS-treated control lymph nodes rarely yielded PCNA-positive cells, and in those rare cases PCNA-positive cells were seen in medullar areas (data not shown). Finally, ConBr, DGL and DVioL were found to increase the ex vivo expression of CD25 on the surface of stimulated lymph node cells (Fig. 3). CD25 comprises the a chain of the IL-2 receptor, and its expression has also been demonstrated to be up-regulated in T lymphocytes upon stimulation with ConA (Black et al. 1988). Cultivation in medium for 24 h in the absence of in vitro stimulus (Fig. 4) demonstrated that cells in stimulated lymph nodes have increased proliferative capacity, as compared to controls (p<0.0001). In both evaluations, no significant differences could be found among the lectins studied (p=0.2619). Apoptotic foci were present in ConBr, DGL and DVioL treated lymph nodes, with typical morphology by electron microscopy (Fig. 2B). The TUNEL assay was performed to allow a better assessment of the amount and overall distribution of the apoptotic foci observed. Stimulated lymph nodes presented many positive foci (Fig. 2C), whereas controls were completely negative (data not shown). The positive foci in the stimulated lymph nodes had the morphologic features typical of apoptosis. They were located in the parafollicular, interfollicular and subcapsular regions, and were seldom present in medullar areas. Focal inflammatory reactions with abundant macrophages and neutrophils were seen in the T lymphocyte areas of the stimulated lymph nodes, especially in DGL and DVioL stimulated lymph nodes (Fig. 2D, and data not shown). Control lymph nodes rarely presented very discrete inflammatory infiltrates (data not shown). Hemorrhagic lymph nodes with associated fibrinoid necrosis of high endothelial venules were frequently observed after stimulation with D. violacea lectin (Fig. 2D). These features were never present in controls or ConBr stimulated nodes and were infrequent in DGL stimulated ones (data not shown). DISCUSSION The present work demonstrates that the lectins studied are able to stimulate in vivo the cells of BALB/c mice popliteal draining lymph nodes. This stimulation is accompanied by the occurrence of apoptosis, inflammation and in some cases necrosis of high endothelial venules. The results presented here also stress the need for investigating possible side effects of lectins with therapeutic possibilities. Lymph node enlargement and increased lymph node cell counts were observed upon lectin treatment, features that had previously been demonstrated following ConA and ConBr administration (Bento et al. 1993). Rodriguez et al. (1992) have reported local increases of mononuclear and polymorphonuclear cells after intra-peritoneal administration of ConA, ConBr and DGL in BALB/c mice, a phenomenon that they have attributed to lectin-induced cellular influx to the site of injection. In our study, the cells in stimulated lymph nodes presented increased IL-2R surface expression and also high proliferation capacity. Abundant lymphoblasts and cells undergoing mitosis, assessed as positivity for PCNA, a cyclin that participates in DNA replication and is present in high amounts from S phase until late G2 (Celis et al. 1988), were seen only in T-cell dependent areas. These findings suggest that during the time period evaluated, the lectins tested were able to stimulate T but not B lymphocytes, and that local activation of T cells probably plays an important role on the observed increases of cell numbers in the draining nodes. The lectins studied were also able of inducing apoptosis in the T-cell dependent areas of stimulated lymph nodes. Lectin-induced T cell activation has been frequently associated with apoptosis occurrence (Kabelitz et al. 1994). Apoptotic cell death can result from the synthesis of potent oxidizing agents, namely nitric oxide (NO), by T cell activated macrophages (Mills 1991, Denham & Rowland 1992). We have previously shown NO production by macrophages induced in vivo upon the intraperitoneal administration of the lectins from C. brasiliensis and D. grandiflora (Andrade et al. 1999). Focal inflammatory reactions were seen in the T lymphocyte areas of the stimulated lymph nodes, especially upon DGL and DVioL administration. The s.c. administration of 50 µg of lectin in the footpads produced hemorrhagic lymph nodes with associated fibrinoid necrosis of high endothelial venules after stimulation with D. violacea lectin, but rarely in DGL stimulated lymph nodes and never in ConBr stimulated ones. Lima et al. (1993) have previously reported strong inflammatory response and leukocyte influx to the site of injection upon subcutaneous administration of high doses of DGL, involving venule necrosis and hemorrhagic appearance. The reaction, similar to the effects of immune complex deposition, may have the participation of TNF-a;, which has been shown to be produced upon in vitro lectin stimulation (Hajto et al. 1990, Amore et al. 1993, Ribereau-Gayon et al. 1997). The three lectins used in this work (ConBr, DGL and DVioL) have the same nominal monosaccharide specificity for Man and Glc residues, but much higher affinities for the branched chain trimannoside, 3,6-di-O-(a-D-mannopyranosyl)-D-mannose (Dam et al. 1998), which is found in the core region of all asparagine-linked carbohydrates. Furthermore, the lectins essentially recognize the same set of binding epitopes on the structure of the trimannoside. The x-ray crystal structures of ConA (Naismith & Field 1996) and DGL (Rozwarski et al. 1998) in complex with the core trimannoside show conserved contact residues for both proteins. The x-ray crystal structure of ConBr in complex with the core trimannoside is not yet determined; however, the alignment of its primary structure with those of ConA and DGL shows that ConBr also has these same contact residues (Fig. 5). Although the complete primary sequence of DVioL is not known, its N-terminal sequence is highly homologous with those above (Calvete et al. 1999), and residues 12-16 that directly bind to the core trimannoside are conserved. However, the differences observed in the intensity of inflammation and associated necrosis suggest subtle variations in the biological activity of these Diocleinae lectins, in spite of their high homology. What characteristics could then account for a differential behavior for the lectins studied? These lectins differ in their binding specificities for complex type carbohydrates (Gupta et al. 1996, Dam et al. 1998). In addition, they show distinct pH-dependent dimer-tetramer equilibrium (Grangeiro et al. 1997, Calvete et al. 1999). Only the tetravalent form is able to cause cross-linking of receptors on the cell membrane, thereby triggering the activating signal transduction pathways. Therefore, the differences in the intensity of inflammation and associated necrosis induced by ConBr, DGL and DVioL can result from small changes of key amino acid residues along the primary structure of the lectins, which affect both the affinity for complex carbohydrate and the ratio between divalent and tetravalent species. ACKNOWLEDGEMENTS To Dr Johan Van Weyenbergh and Dr Tânia Correia for helpful suggestions, and to Mr Jorge Tolentino, Mrs Maria Lúcia Vieira and Mrs Cristina dos Santos Mota for technical assistance. REFERENCES
This work was supported by PADCT (grant number PADCT02-5BIO-01/97-02/01-47). TB and SA received fellowships from Brazilian National Research Council (CNPq). BC, TBG, LARF and MB-N are senior investigators from CNPq. Copyright 2001 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc01105f5.jpg] [oc01105f4.jpg] [oc01105f2.jpg] [oc01105f1.jpg] [oc01105f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}