 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(5) July 2001, pp. 709-712 Variants of the Plasmodium vivax Circumsporozoite Protein (VK210 and VK247) in Colombian Isolates JM González+, S Hurtado, M Arévalo-Herrera, S Herrera Instituto de Inmunología
del Valle, Universidad del Valle, Carrera 4B, No. 43-00, Cali, Colombia Received 5 April 2000
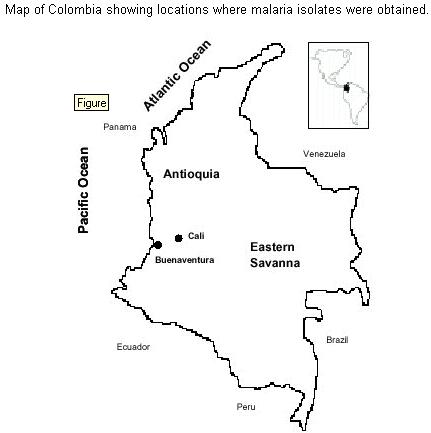
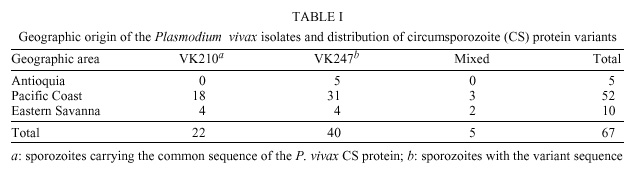
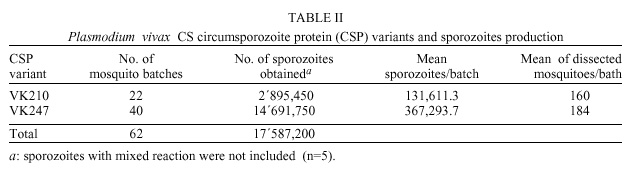
Code Number: oc01112 Phenotypic diversity has been described in the central repeated region of the circumsporozoite protein (CSP) from Plasmodium vivax. Two sequences VK210 (common) and VK247 (variant) have been found widely distributed in P. vivax isolates from several malaria endemic areas around the world. A third protein variant called P. vivax-like showing a sequence similar to the simian parasite P. simio-ovale has also been described. Here, using an immunofluorescent test and specific monoclonal antibodies, we assessed the presence of two of these protein variants (VK210 and VK247) in laboratory produced sporozoite. Both sequences were found in parasite isolates coming from different geographic regions of Colombia. Interestingly, sporozoites carrying the VK247 sequence were more frequently produced in Anopheles albimanus than sporozoites with the VK210 sequence. This difference in sporozoites production was statistically significant (p <0.05, Kruskal-Wallis); not correlation was found with parameters as the total number of parasites or gametocytes in blood from human donors used to feed mosquitoes. Previous studies in the same region have shown a higher prevalence of anti-VK210 antibodies which in theory may suggest their role in blocking the development of sporozoites carrying the CSP VK210 sequence. Key words: Plasmodium vivax - sporozoites - Anopheles - Colombia The circumsporozoite protein (CSP) is the major coat protein of the malaria sporozoites. It presents a central repeat domain flanked by non-repeated amino and carboxyl sequences containing highly conserved protein stretches (regions I and II). These flanking regions also display some degree of polymorphysm (Mann et al. 1994). In Plasmodium vivax the first sequenced gene encoded a tandemly repeated nonapeptide GDRA(D/A)GQPA that later was termed common or VK210 sequence (Arnot et al. 1985). Phenotypic heterogeneity in the repeated region of the P. vivax CSP was initially described in parasites isolates from Thailand expressing the nonapeptide ANGA(G/D)/N/D)QPG, termed variant or VK247 sequence (Rosenberg et al. 1989). A third sequence from the CSP was obtained from a parasite resembling morphologically to P. vivax. The CSP repeat sequence (APGANQEGGAA) was found to be similar to the simian malaria parasite P. simio-ovale (Qari et al. 1993a). Serologic surveys in several P. vivax endemic areas demonstrated the presence of antibodies that recognize recombinant and synthetic polypeptides that represent the three CSP variants (Wirtz et al. 1990, Franke et al. 1992, Marrelli et al. 1998). Using DNA techniques (Kain et al. 1991), the VK210 and VK247 sequences were found in P. vivax isolates distributed worldwide (Mann et al. 1994, Sattabongkot et al. 1994). Despite the fact that this third CS sequence was found in parasite isolates around the world (Qari et al. 1993b), its global distribution was not confirmed in another study (Gopinath et al. 1994). Wild parasite isolates display a number of silent mutations throughout the whole CSP gene sequence, these mutations do not represent changes in the protein sequence (Mann et al. 1994). Here, we report the frequence of the two CSP variants (VK210 and VK247) in wild P. vivax isolates from Colombia as determined by direct immunofluorescent test (IFAT) in laboratory produced sporozoites. We also describe the susceptibility of Anopheles mosquitoes to the experimental infection with parasites carrying one of these two CSP sequences. Human blood samples were obtained from 70 individuals who requested medical attention at the Malaria Eradication Service (SEM) in Cali and Buenaventura during 1994-1996. These individuals came from three different geographic regions of Colombia: Antioquia, the Eastern Savanna and the Pacific Coast (Figure). Infection by P. vivax parasite was diagnosed in all patients using Giemsa stained thick and thin smears. Whole blood samples were obtained by venous puncture using heparinized syringes and transported to the laboratory within 1 h after bleeding. Adult female An. albimanus mosquitoes from two different strains: Tecojate (Guatemala) and Buenaventura (Colombian Pacific Coast) were used to produce sporozoites. Batches of 500 mosquitoes were put in individual cages and fed using artificial membrane feeders containing the P. vivax infected human blood. Mosquitoes were then kept at 27ºC and 85% of relative humidity. Infection was confirmed at day 8 after blood feeding by the observation of gut oocysts in a sample of exposed mosquitoes. Salivary glands were dissected in sterile PBS 1x pH 7.4 at day 14 after infection and the number of sporozoites was calculated using a blood count chamber (Salas et al. 1994). Slides for IFAT were prepared using air-dried sporozoites. Two mAbs (kindly provided for R Nussenzweig, New York University) specific to P. vivax CSP were used: 2F2 and 2E10 which recognized VK210 and VK247, respectively. A third mAb 2A10 which recognized the central region of P. falciparum was used as a negative control. Reaction was revealed using 1/50 PBS diluted IgG anti-mouse FITC (Sigma). Evaluation was carried out by two independent readers with a fluorescent light microscope with 40x and 100x immersion oil lenses. Each isolate was tested at least twice in duplicate or triplicate spots. Information obtained was processed and analyzed using the EPINFO software. A total of 67 P. vivax sporozoite isolates representing 96% of blood samples used for mosquito feeding were analyzed. The isolates were from The Pacific Coast region 52 (77.6%), The Eastern Savanna region 10 (14.9%) and Antioquia 5 (7.5%); these three regions contributing yearly with 87% of the malaria cases in Colombia. IFAT showed the VK210 sequence in 22 (32.8%) isolates an the VK247 sequence in 40 (59.7%) isolates. Five (7.5%) of the samples reacted with both mAbs indicating the presence of mixed infection (Table I). Most sporozoite lots (60/90%) were produced in An. albimanus Tecojate strain and the remaining (7/10%) in Buenaventura strain. All sporozoites obtained in the Buenaventura strain displayed the VK247 sequence. Five isolates containing mixed reaction were obtained in the Tecojate strain. There was a significant statistical correlation between the phenotype of the central region of the P. vivax CS protein and number of sporozoites obtained (p <0.05, Kruskal-Wallis). As shown in Table II, sporozoites with VK247 were produced more abundantly than VK210. There was not statistical correlation between other variables such as total parasitaemia or gametocitaemia in the blood donors and number of sporozoites produced. In a recent study, it was demonstrated a differential susceptibility of Anopheles mosquitoes to malaria infection with P. vivax isolates in Mexico. Indeed, An. albimanus and An. pseudopunctipennis were preferentially infected by VK210 and VK247 variants, respectively (González-Ceron et al. 1999). In our work, comparison of sporozoite rates between the two Anopheles strains used (Tecojate and Buenaventura) are not possible due to differences in the number of exposed batches. Naturally acquired antibodies that recognize the VK247 sequence have been demonstrated less prevalent than those recognizing VK210 in several countries: Thailand (Wirtz et al. 1990), Peru (Franke et al. 1992), Brazil and India (Cochrane et al. 1990). Malaria cases in Colombia for the period of this study (1994-1996) were predominantly due to P. vivax (62%). We have previously demonstrated that anti-CSP antibodies in donors from the Colombian Pacific Coast to VK210 are more frequent (68-75%) than to VK247 (11-20%) (Arévalo-Herrera et al. 1998). However, we report here a predominance of malaria isolates carrying the VK247 sequence (67%) when the used read-out is the sporozoite production. Putting together our data it appears as if anti-CSP antibodies against VK210, which are more prevalent in the same geographic region, could block the development of the VK210 sporozoites and therefore they become less frequent in mosquitoes. Alternatively, it might be that VK210 is more immunogenic that VK247 and that would explain that lower prevalence of VK210 sporozoites induces higher antibody prevalence. This might result in the selection of the VK247 sporozoites. It is clear that CSP is required to complete the cycle inside the mosquito (Menard et al. 1997). In contrast to this hypothesis, previous reports using P. falciparum showed enhancing of sporozoite production when antibodies to the central region were present in the serum (Do Rosario et al. 1989). Current experiments addressing this question are going on. These findings will allow us to continue expanding our knowledge on the behavior of P. vivax CSP and to understand the complex relationship between parasites, vectors and the human immune response. ACKNOWLEDGMENTS To M Salas, H Ortiz , JC Millán and MF Suárez for their help and technical assistance. REFERENCES
This work was supported by the Fondo Colombiano de Investigaciones Científicas y Proyectos Especiales" Francisco José de Caldas" (Colciencias). Copyright 2001 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc01112f1.jpg] [oc01112t2.jpg] [oc01112t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}