 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(s.1) 2001, pp. 79-83 Preliminary Analysis of Sm14 in Distinct Fractions of Schistosoma mansoni Adult Worm Extract Nilton Thaumaturgo, Mônica Magno Vilar, Catia Maria Diogo, Ricardo Edelenyi, Miriam Tendler+ Laboratório de Esquistossomose Experimental,
Departamento de Helmintologia, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365,
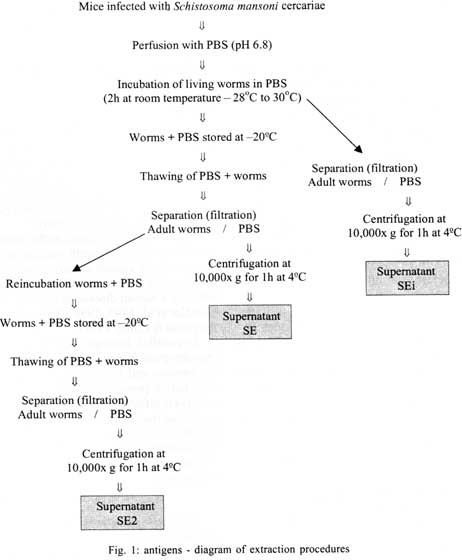
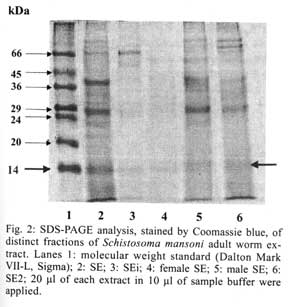
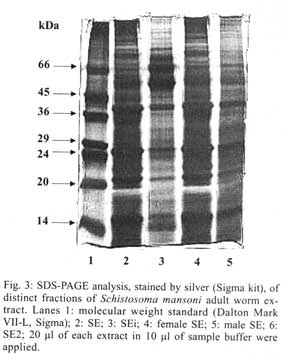
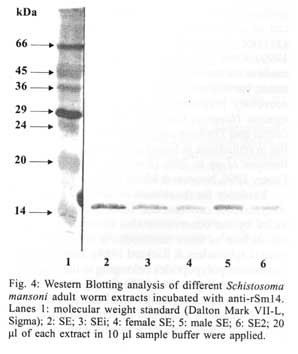
21045-900 Rio de Janeiro, RJ, Brasil Received 14 May 2001 Code Number: oc01155 In previous studies it was shown that the recombinant molecule, r-Sm14, induces high levels of protection against Schistosoma mansoni infection in two outbred animal models and immune crossprotection against infection by Fasciola hepatica in Swiss outbred mice. r-Sm14 was derived from a living worm extract, called SE, and is being developed as the molecular basis of an anti-helminth bivalent vaccine against the two parasites, for medical and veterinary application. Present data refer to SDS-PAGE and Western Blotting analysis of four different preparations of S. mansoni adult worms focusing Sm14 identification. The extracts correspond to the initial fraction of the SE extraction process, containing products released by living worms (SEi); SE2, reextraction of adult worms in PBS; and SE of separated male and female adult worms. In all extracts it was possible to detect the component of 14 kDa, that was recognized by specific anti-rSm14 antibody raised in rabbits. Key words: Schistosoma mansoni - vaccine - excretory/secretory antigens - FABP - Sm14 Schistosomiasis is the second most prevalent tropical disease in the world – after malaria – with about 200 million adults and children infected in 74 countries. It is estimated that 20 million of them have a serious form of the disease or related disability, and that 200,000 people die from the disease every year (WHO 1996). The magnitude of the endemy fully justifies the interest for the development of effective methodology for transmission control, based on an anti-schistosomiasis vaccine. In addition, reinfection after treatment, particularly in young inhabitants of endemic areas, makes chemotherapy expensive. The available drugs have no effect on already developed hepatosplenic manifestations and do not affect transmission rates. Naturally acquired partial immunity is well demonstrated in endemic areas, with reduced intensity of infection in adolescents and older age groups as compared to children. Previous studies in our laboratory showed that vaccination of Swiss outbred mice and New Zealand rabbits with a saline extract of adult Schistosoma mansoni worms (SE), resulted in 70% and 90% of protection respectively, against cercarial challenge (Tendler et al. 1986, 1991). Focusing the molecular characterization of SE components, towards the goal of identifying the immunoprotective components, it was obtained by gene cloning a 14 kDa molecule, named Sm14, belonging to family of fatty-acid binding proteins – FABPs (Moser et al. 1991, Tendler et al. 1996). It was demonstrated that Sm14 can reproduce the same high protection levels of SE against S. mansoni experimental infection and also be protective against another digenetic trematode, Fasciola hepatica (Tendler et al. 1995, 1996). Sm14 is thus considered as the molecular basis of a bivalent anti-helminth vaccine, aimed for the veterinary use against an economically important disease such as fascioliasis, and schistosomiasis which is a human disease of developing countries (Tendler et al. 1995, 1996, Bergquist 1995, 1997, Bergquist & Colley 1998). In parallel, biological parameters other than immuno-protection, related to Sm14 localization in the parasite and function, are being studied. Results herein presented aimed at the investigation of Sm14 in different adult worm extracts, corresponding to four fractions of SE. SEi, initial fraction of the SE extraction process where worms are still alive; SE2, with products obtained by saline reextraction of adult worms; and SE of separated male and female worms. MATERIALS AND METHODS Parasites - LE strain (Belo Horizonte, Brazil) of S. mansoni is maintained in our laboratory by passage through Swiss mice and Biomphalaria glabrata. Parasites were obtained by retrograde perfusion with heparinized saline (0.85% NaCl solution), 45 days post-infection (Pellegrino & Siqueira 1956), and used for preparing the different extracts, as described below. Preparation of adult-worm-derived extract (SE) - Parasites were rinsed in PBS. Living worms were then allowed to stand at room temperature (28°C to 30°C) in fresh PBS (phosphate buffered saline) for 2-3 h, and frozen at -20°C. After thawing, PBS suspensions of the worms (1g worms/10 ml PBS) were filtered through a wire mesh, and centrifuged at 10,000 g for 1 h at 4°C (Tendler & Scapin 1981, Tendler et al. 1982). The protein content of SE was assessed by Lowry's method (Lowry et al. 1951), and stock batches containing 1 mg/ml were stored at -20°C. Male and female extracts were prepared according to the same methodology, with parasites separated immediately after perfusion. Preparation of a rapidly released "fraction" of adult-worm-derived extract (SEi) - SEi preparation corresponded to the initial step of SE: after perfusion as described for SE, live worms were incubated for 2-3 h at room temperature (28oC to 30oC), in PBS. Thereafter, adult worms were filtered from incubation solution (SEi) in a wire mesh. Preparation of SE2 - Adult worms remained from SE initial preparation, were re-stored frozen (-20°C) in PBS for one week (1g worms/10 ml PBS). After thawing, PBS suspensions were filtered through a wire mesh, and centrifuged at 10,000 g for 1 h at 4°C. Preparation of antigens as above, are schematically represented in Fig. 1. Antibody - Monoespecific policlonal antibody, anti-rSm14, was obtained in male New Zealand rabbits immunized with two footpads injections of 50 µg r-Sm14 in Freund's complete adjuvant each, administrated one week apart, followed by an intraperitoneal booster of 50 µg r-Sm14, 21 days after the second injection. Animals were bled for serum collection at intervals of 20 days, starting from pre-immunization up to pre-challenge date, at the peripheral ear vein. SDS-PAGE and Western Blotting analysis - Twenty µl per lane of SE fractions (SEi, SE2, SE male and SE female) were loaded on 15% SDS-PAGE. Gels were either stained with Coomassie blue, or transferred into nitrocellulose membrane according to the method originally reported by Towbin et al. (1979). Nitrocellulose membranes were blocked using a nonfat dry milk solution (5% in PBS). Nitrocellulose strips were then washed in PBS/Tween, and subjected to incubation with specific anti-rSm14 antibody (diluted 1:1,000) for 2 h. Following extensive PBS/Tween washing, appropriate alkaline phosphate-coupled goat anti-rabbit antibody (diluted 1:5,000) were applied for 2 h. Filters were again washed in PBS/Tween, and color reaction was developed with NBT (nitro-blue-tetrazolium) and BCIP (5-bromo-4-chloro-3-indolyl phosphate) both at 50 mg/ml. RESULTS In all tested extracts male and female S. mansoni saline extract (SE), initial fraction of the SE extraction process were worms are still alive (SEi) and saline reextraction of adult worms (SE2), it was possible to detect, by 15% SDS-PAGE stained by silver, a 14 kDa band that comigrated with purified a-lactalbumin from SDS-PAGE standards (Figs 2, 3). As shown in Western blotting analysis, the 14 kDa component, identified in the four preparations, was recognized by monospecific policlonal anti-Sm14 antibody raised in New Zealand rabbits (Fig. 4). DISCUSSION Products released by worms as excretory/secretory antigens have been considered as important source of antigens capable of generating immune response in hosts (Maizels et al. 1982, Lightowlers & Rickard 1988, Maizels & Selkirk 1988, Lightowlers 1996, Wilson & Coulson 1998, Maizels et al. 1999). Some excretory/secretory products are clearly associated with specialized excretory or secretory organs. However, molecules derived from the tegument or cuticle of helminths make a substantial contribution to the contents of excretory/secretory products of the parasite maintained in vitro, and are probably also released in vivo (Lightowlers & Rickard 1988). The active release of antigens from the surface of helminths should have a potentially significant impact on the immunobiology of the helminth-host relationship throught their potential triggering effect on the immune system of infected hosts (Kaushal et al. 1982, Maizels et al. 1982, Butterworth 1984, Lightowlers & Rickard 1988, Matsumoto et al. 1988). Since 1955, worms excretory/secretory antigens have been used in immunization experiments, as shown in mice immunized with Trichinella spiralis excretory/secretory products, where the parasites were expelled more rapidly in vaccinated animals than in control group (Campbell 1955). Subsequent testing in a wide range of parasitic infections has proven excretory/secretory proteins to be successful immunogens (Lightowlers & Rickard 1988). Natural immunity to gastro-intestinal nematodes will develop in older animals and, with continued exposure to infective larvae, the immunity is maintained. This protective response can largely be attributed to immune response to excretory/secretory antigens (Newton & Munn 1999). Over the years, many attempts have been made to induce immunity to gastro-intestinal nematodes, particularly those of sheep, with excretory/secretory preparations, including vaccination against Haemonchus contortus, Taenia circum-cincta and Trichostrongylus colubriformis, resulting in reductions in faecal egg counts and/or worm burdens of up to 70% (Emery & Wagland 1991, Emery 1996, Newton & Munn 1999). Evidence for the release of surface antigens of Ancylostoma caninum and Toxocara canis is provided by the observation that antibody bound to the surface of these nematodes is shed rapidly in vitro (Lightowlers & Rickard 1988). Moreover, recombinant polypeptides belonging to the Ancylostoma secreted protein (ASP) family have shown promise for reducing hookworm burdens (50-80%) after A. caninum challenge infection in mice. The ASP family members have been found in a wide variety of nematodes including A. duodenale and Necator americanus (Hotez et al. 1999). In vitro release of surface-derived antigens as excretory/secretory products was shown in filarial nematodes, such as adult Brugia malayi (Kaushal et al. 1982) and B. pahangi (Maizels et al. 1985). S. mansoni infection, it was demonstrated that surface components of S. mansoni schistosomula and adult worms are shed in vitro and probably also in vivo (Kusel & Mackenzie 1975, Kusel et al. 1975). Matsumoto et al. (1988) suggested that paramyosin may be also an unexpected secretory product of S. mansoni, and related the localization of this protein to the tegument of the parasite with the basis for resistance observed in mice immunized with paramyosin. Similarly to schistosomes, the structure of F. hepatica tegument suggests active secretion of material at the parasite surface (Hanna 1980). Another molecule, Sm62, a citokeratin-like protein, was detected as a secretory unexpected product (Diogo et al. 1994). Results herein presented were derived from the investigation of the S. mansoni FABP, Sm14, in adult saline extracts (SE), designed to contain excreted/secreted and somatic antigens: SEi – corresponding to components early released by living adult worms (excretory/secretory antigens) and SE2 – containing components released by previously freezed/thawed adult worms. SDS-PAGE and Western blotting analyses allowed the detection of a 14 kDa component, that was recognized by specific anti-Sm14 immunosera, in SEi and SE2, with similar amounts of proteins as indicated by SDS gel patterns. Extracts of separated male and female adult worms, were also utilized for the characterization of putatively sex related protective antigens. In fact, Sm14 was detected in both preparations, and this fraction was further recognized, by specific anti-rSm14 immunosera, in immunoblotting. In addition, the 14 kDa band was stronger in male extract than in the female, suggesting that Sm14 may be constitutively present and/or excreted/secreted in higher amounts in male adult worms. ACKNOWLEDGEMENT To Dr Roberto Magalhães Pinto for critical review of the manuscript and valuable comments. REFERENCES
This work was supported by Fiocruz, Faperj and CNPq. Copyright 2001 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc01155f2.jpg] [oc01155f4.jpg] [oc01155f3.jpg] [oc01155f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}