 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 96(7) 2001, pp. 889-894 Adult and Nymphs of Microtriatoma trinidadensis (Lent, 1951) (Hemiptera: Reduviidae) Caught from Peridomestic Environment in Bolivia J De la Riva/+, A Matias, M Torrez, E Martínez, JP Dujardin* Instituto Nacional de Laboratorios en Salud,
Rafael Zubieta N° 1889, La Paz, Bolivia *Unité Mixte de Recherche IRD-CNRS
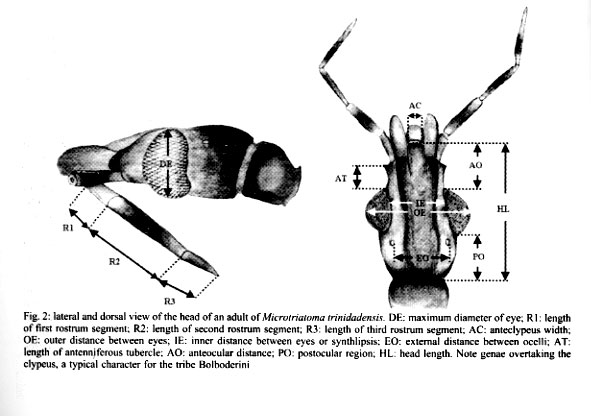
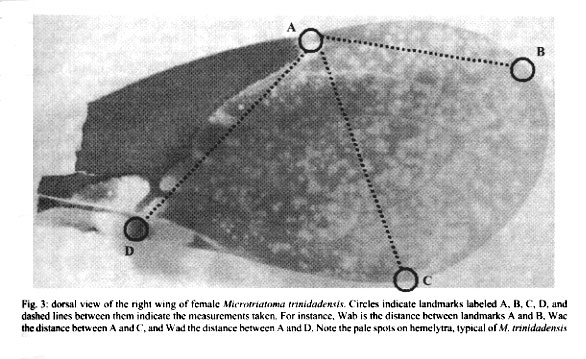
9926, Montpellier, France Received 17 July 2000 Code Number: oc01175 This is the first report of adult and nymphs (20 nymphs of all stages and 4 adults) of Microtriatoma trinidadensis (Lent 1951) (Hemiptera: Reduviidae: Triatominae) collected in peridomestic environment, in the department of La Paz, Bolivia. These specimens were associated to Rhodnius stali Lent, Jurberg & Galvão 1993. The exceptional finding of M. trinidadensis in peridomestic environment, illustrates the general tendency of triatominae to adapt to human dwellings and dependences. Key words: Microtriatoma trinidadensis - Rhodnius stali - peridomestic - coprophagy - Bolivia Microtriatoma mansosotoi Prosen & Martínez 1952 was described by Carcavallo et al. (1976) and synonymized later with M. trinidadensis Lent, 1951 by Lent and Wygodzinsky (1979), who described adults and fifth nymph only, inferring from them the characteristics of the remaining stages. The first specimens of Microtriatoma reported in Bolivia as M. mansosotoi Prosen & Martínez 1952 were collected in the Chapare (Department of Cochabamba, Bolivia) (Torrico 1958) and in Santa Cruz (Department of Santa Cruz, Bolivia) at the periurban zone (Wygodzinsky 1958). Since then, Microtriatoma was mentioned many times in South America (Venezuela, Colombia, Surinam, Peru, Bolivia and Brazil) (Lent & Wygodzinsky 1979, Miles et al. 1981, Pinto 1992), whereas it was reported only a few times in Central America (Trinidad and Panama) (Carcavallo et al. 1999). This species is known to be associated with palms and hollow trees, hidden in tree cavities such as folded leaves, especially within nests of Didelphis marsupialis (Lent & Wygodzinsky 1979, Miles et al. 1981). According to Miles et al. (1981), M. trinidadensis could feed on opossum in the wild. Under laboratory conditions it could feed readily on mice at least during some time, after what it died. M. trinidadensis has been found naturally infected by Trypanosoma cruzi (Miles et al. 1981, Silveira 1998). This work describes for first time the presence of M. trinidadensis in peridomestic environment in La Paz Department, Bolivia (Fig. 1) associated with Rhodnius stali Lent, Jurberg and Galvão, 1993 and provides a morphological description of all nymphal instars. MATERIALS AND METHODS Field survey - During May 1998, we carried out a field survey aiming to collect any Triatominae in domestic and peridomestic environment in Alto Beni, a sub-Andean region of the Department of La Paz (Fig. 1) at the border with Amazonian region (15°19' S, 67°32' W). It has an annual mean temperature of 25.89°C and a mean relative humidity of 80.6%. Insects Morphological examination - The external morphological examination and classic taxonomic identification was performed without dissection of genitalia, using as reference the description of Lent and Wygodzinsky (1979). Only specimens identified as M. trinidadensis Lent 1951 were included in the subsequent studies. Eleven nymphs of this species were stored at -70°C for further studies and the remaining 9 were included in the INLASA collection. Two adults were sent later to the reference laboratory of Instituto Oswaldo Cruz (IOC), Rio de Janeiro, Brazil for morphological confirmation and deposited by Carcavallo at the IOC collection. Parasite search - Rectal contents of 3 adults and 3 fifth instar nymphs were examined microscopically for the presence of flagellates. Morphometrics - Using a monocular micrometer eyepiece (10x), we performed 12 head and 3 wing measurements in 2 adult specimens (1 male and 1 female). For comparison of the specimens collected with known material, 2 other adults were measured from the Carcavallo collection at IOC: 1 type female of M. trinidadensis (named here F2), and 1 allotype female of M. mansosotoi (named here F1). Head measurements (Fig. 2) - Maximum diameter of eye (DE); length of first rostrum segment (R1); lenght of second rostrum segment (R2); lenght of third rostrum segment (R3); anteclypeus width (AC); outer distance between eyes (OE); inner distance between eyes or synthlipsis (IE); external distance between ocelli (EO); lenght of antenniferous tubercle (AT); anteocular distance (AO); postocular region (PO); head length (HL). The length of antenniferous tubercle is an oblique measure, not parallel to the head length measurement. Wing measurements were the linear distance between the points indicated in Fig. 3: Wab, distance between A and B; Wac, distance between A and C; and Wad, distance between A and D. RESULTS Insects - A total of 24 specimens collected were identified as M. trinidadensis, 20 nymphs: 2 first instar (N1), 2 second instar (N2), 1 third instar (N3), 3 fourth instar (N4), 12 fifth instar (N5); and 4 adults (2 females and 2 males). All the 24 specimens came from one house only, out of the 101 houses examined. The specimens collected were found in a chicken coop built with material from the palm tree Attalea phalerata. It is worth mentioning that chicken coops were not frequent in this area (there were 20 of them for 101 houses), and that the other chicken coops were generally made of different materials (adobe for instance). Another particularity was the joint finding, at the same place, of a few Rhodnius stali together with M. trinidadensis (Matias et al. 2001). In the laboratory, various potential hosts (chicken, human, mice) were proposed to allow the insects to feed, without success. The unique feeding behavior observed was a fifth instar nymph probing feces of R. stali, which apparently could trigger its last molting. Parasite search - No flagellate was observed in the fecal drops. Morphological and morphometric description - As hypothesized by Lent and Wygodzinsky (1979), all nymphs presented spongy fossulae on the 3 pairs of legs. In both adults and nymphs, femur and tibia were without spines or denticles on under surface, and proximal and distal extremities of each segment of the legs were generally darker. Thorax was not granulose, with numerous short scale-like setae. The connexivum patterns on the first (N1) and second instar nymph (N2) were poorly noticeable, and became more conspicuous on the following stages (N3, etc.). The overall color of N1 and N2 was tenuous rose and maintained so during their lifetime (two weeks). This color of nymphs abruptly changed from tenuous rose in the first instars (N1, N2) to obscure brown in the next ones (N3, N4, N5 and adults). Another feature changing between N2 and N3 was the relative lengths of head and rostrum. In the first nymphs (N1, N2), the rostrum reached the neck. From the third instar nymph to the following stages, the size of second rostrum segment (R2) grew up so that the complete rostrum presented larger dimensions than the length of the head. Our measurements (Table) were compatible with the dimensions of two other specimens of Microtria-toma conserved as type and allotype in the Carcavallo collection at IOC: one female of M. mansosotoi (F1, Table), and one female of M. trinidadensis (F2, Table). DISCUSSION This is the first report of M. trinidadensis in the peridomestic environment. The main morphological characteristics of our sample were as described by Lent and Wygodzinsky (1979). We could confirm the presence of spongy fossulae on the 3 legs of all instars, and observe some abrupt morphological changes between second and third instar nymphs: the color, the visibility of con-nexivum spot (Fig. 4b) and relative proportions of head and rostrum. Interestingly, our measurements reproduced almost exactly the dimensions of M. mansosotoi female allotype, as identified in Cochabamba (Villa General Román, Chapare, Bolivia) by Torrico (1958). This could suggest that the Bolivian population of M. trinidadensis could represent a slight but stable geographic variation from the M. trinidadensis holotype. The present finding adds a third Department of Bolivia (La Paz) to the Bolivian distribution of M. trinidadensis. The presence of M. trinidadensis just in one dwelling, out of 101 checked, could appear as an accident. However, not the 101 dwellings had chicken coops: 20 chicken coops were examined, of which 5 were positive for the presence of R. stali Lent, Jurberg and Galvão, 1993 (Dujardin et al. 2000), and M. trinidadensis was found in 1 of these 5 chicken coops. The presence of nymphs of all instars suggests that M. trinidadensis was breeding there. The apparent absence of eggs was probably due to their very small size (0.8 mm of half diameter and half length 1.1 mm as described by Barata (1995), making them almost invisible. Our search for flagellates was negative. This does not discard the possibility of infection by T. cruzi. For the specimens collected in this survey, the probable source of food was hens, which, like birds in general, are known to be refractory to the T. cruzi infection. In other associations (as with opossum), M. trinidadensis has been found infected by T. cruzi (Miles et al. 1981, Silveira 1998). These specimens were collected in a chicken coop which was also colonized by R. stali, a similar association has been reported before with R. robustus Larrouse 1927 in Venezuela (Lent & Wygodzinsky 1979). These references and our observation of a M. trinidadensis nymph probing feces of R. stali could represent some kind of predatory behavior and could reflect a true biological association. It also could provide a first explanation to the presence of M. trinidadensis in peridomestic structures. In support of this, Belminus peruvianus Herrer, Lent and Wygodzinsky, 1954 (another Bolboderini), was found to colonize houses in the Marañon Valley of Peru, where it was observed to attack the Chagas disease vector Panstrongylus herreri Wygodzinsky 1948 (Herrer 1955). Similarly, cleptohematophagy of B. herreri Lent & Wygodzinsky, 1979 on specimens of R. prolixus Stål 1859 has been described in Colombia (Sandoval et al. 2000). Whatever the mechanism explaining this unusual ecotope for M. trinidadensis, our finding illustrates again the general tendency of Triatominae to adapt to domestic structures (Dujardin et al. 2000). As long as the hypothesis is true of a biological association between R. stali and M. trinidadensis, in a geographic area where R. stali is known to adapt to domestic structures (Dujardin et al. 2000, Matias et al. 2001), M. trinidadensis could be just following its source of food. REFERENCES
This work has benefited from international collaboration through the ECLAT network. Copyright 2001 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc01175t1.jpg] [oc01175f1.jpg] [oc01175f2.jpg] [oc01175f4.jpg] [oc01175f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}