 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(7) 2001, pp. 947-950 SHORT COMMUNICATION Rhodnius robustus in Bolivia Identified by its Wings A Matias, JX de la Riva, M Torrez, JP Dujardin*/+ INLASA, La Paz, Bolivia *UMR IRD-CNRS 9926 UR
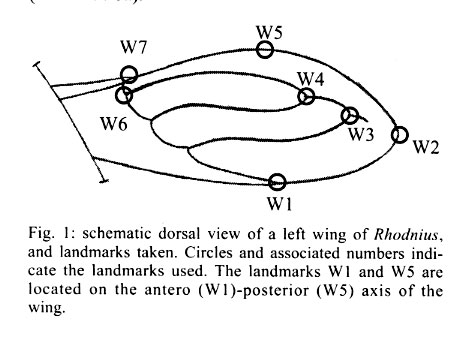
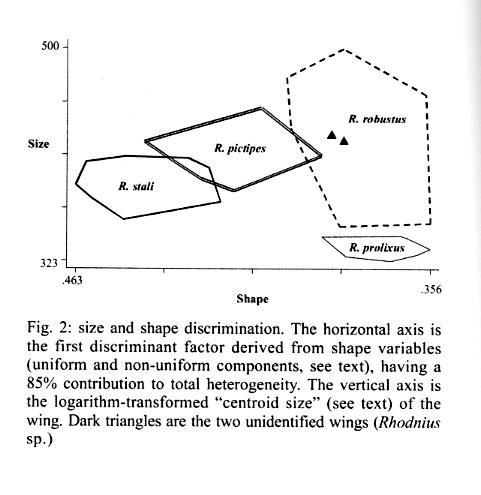
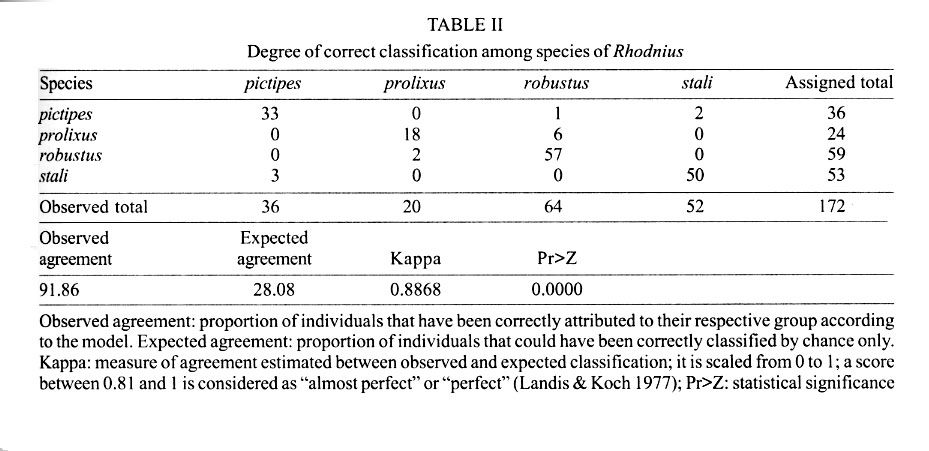
062, IRD La Paz, La Paz, Bolivia Received 4 December 2000 Code Number: oc01184 Wings of a Rhodnius specimen from Alto Beni (Bolivia) was examined for identification and compared with R. stali, R. robustus, (certified Bolivian species), R. pictipes and R. prolixus (suspected Bolivian species). A projection of the unidentified wings as supplementary data into a discriminant analysis of shape revealed clear cut differences with R. stali and R. pictipes, less differences with R. prolixus, and none with R. robustus. Combining global size and shape of the wings, the unknown specimen was identified as R. robustus. Thus, this study confirmed the presence of R. robustus in Bolivia. It also highlighted the possibility of morphometrics to taxonomically interpret one individual, or even one piece of an individual, when related species data are available for comparison. Key words: Rhodnius robustus - Triatominae - geometric morphometry - discriminant analysis - Bolivia Remnant pieces of a dead Rhodnius specimen found in Alto Beni (Bolivia) were examined for species identification. It was compared with R. stali and R. robustus, which are the only certified species of Rhodnius in Bolivia, as well as with two other species which are suspected to be present in Bolivia: R. pictipes and R. prolixus. Only the wings allowed a reliable morphological comparison. Using geometric techniques of morphometrics, the size and shape of these unidentified wings appeared to conform with R. robustus. The first reports of Rhodnius species in Bolivia are those of Torrico (1946, 1958), where only R. pictipes was mentioned. This species has possibly been confounded with R. stali, since no specimen could be further confirmed as R. pictipes. The only published report of R. pictipes after those of Torrico (Tibayrenc & Le Pont 1984) is probably also a misidentification with R. stali (Dujardin et al. 2000). After 1959, R. robustus was reported in the Departments of Pando and Santa Cruz (Wygodzinsky 1959, Lent & Jurberg 1969, Bermudez 1994, Galindez et al. 1996), as well as R. prolixus (Lent & Jurberg 1969, Romero & Borda 1979, Borda 1979). No precise geographic origin was described for R. prolixus, which may be considered as a doubtful report (Bermudez 1994), possibly a confusion with R. robustus. No details were given about relevant, diagnostic morphological traits of Bolivian R. robustus except in the revision of Lent and Jurberg (1969). Unfortunately, none of these specimens was conserved in Bolivia or was presently available for direct comparison. The use of morphometrics on many measurements and of subsequent multivariate analyses for species identification in Triatominae was introduced for the first time by Gorla et al. (1993). The specimen we examined was made of bits and pieces of a dead female in bad morphological conditions of preservation. The general colors and dimension patterns of the head were recognizable, though not usable for a reliable metric analysis, whereas the two wings were perfectly preserved from any mechanical or parasitic damage. It was thus possible to compare this specimen with other species of Rhodnius on the basis of wing traits. We examined its geometry in relation to seven landmarks (Fig. 1). Instead of measurements of distances between landmarks, the geometric method uses coordinates of landmarks (Rohlf & Marcus 1993). This method takes into account the spatial relationship among variables, so that it is expected to have greater statistical power than methods derived from simple distance measurements. By constructing separate variables for general size and for shape variation, the geometric method analyzes the metric differences in term of size-free and size-dependent (allometry) variation. A total of 87 insects (174 wings) were mounted on microscope slides using Hoyer medium (for samples size and respective origins, see Table I). Camera lucida drawings of the wings were made on a microscope at a magnification that allowed maintenance of a consistent plane of focus to control distortion. On the membranous part of the hemelytra, we identified seven landmarks: five of "type I", denoted W1, W3, W4, W6, W7 (tissue intersections), and two of "type II", denoted W2 and W5 (Bookstein 1991) (Fig. 1). The coordinates of landmarks were digitized using TPSdig version 1.15 (Rohlf 1998a). For estimating the global size of the wing, we used the isometric estimator known as "centroid size", derived from coordinate data. It is defined as the square root of the sum of the squared distances between the center of the object and its landmarks and was obtained by using the TPSregr software (TPSregr version 1.15 Rohlf 1998c). The unidentified wings showed one of the largest sizes, compatible with either R. robustus or large R. pictipes, but consistently larger than R. stali or R. prolixus (see the vertical axis of Fig. 2). This later species presented the smallest wings, as already observed before for specimens originating from Honduras (Dujardin et al. 1998). Shape variables, i.e. variables scaled for centroid size, were obtained after Procrustean superimposition of raw coordinates (Rohlf 1996), which were subjected to a "Thin Plate Spline" analysis (Bookstein 1991) using the TPSrelw version 1.17 software (Rohlf 1998b). This later analysis provides parameters subdivided into "uniform" and "non-uniform" components of shape changes (Bookstein 1991). Using both components as input, a discriminant analysis was performed with R. stali, R. pictipes, R. robustus and R. prolixus as groups, but excluding the unknown specimen. This discriminant analysis could satisfactorily classify most of the individuals (the agreement was considered "almost perfect", see Table II). The unidentified wings were then projected on the first discriminant factor as supplementary data (JMP®, SAS Institute Inc. 1995) and their respective positions examined in the scatter plot (STATA®, Computing Resource Center 1992) including size variation. They clearly matched the R. robustus wings (Fig. 2). Since shape was obtained by elimination of the "centroid size" which is an isometric parameter, the resulting superimposed shape coordinates included any allometries. Linear regression of the first shape discriminant factor on isometric size revealed indeed a still significant allometric content (P = 0.002), with however a low explained variance (4%, detailed results not shown). This ensured that the relationships described by our discriminant analysis (Fig. 2) were poorly affected by growth patterns (size variation). Pure shape similarity of the wings of one individual with those of R. robustus is not proof that the questioned specimen is actually R. robustus. At least, it strongly suggests that it belongs to this group of Rhodnius (the prolixus group), which is evolutionary far from the remaining Rhodnius species, either R. stali, R. pictipes or the other ones. Indeed, the Rhodnius genus may be subdivided into a minimum of three distinct clades. The prolixus group (robustus, prolixus, nasutus, neglectus and domesticus), the pallescens group (ecuadoriensis, colombiensis, pallescens) and the other remaining six species among which the closely related R. stali and R. pictipes (Chavez et al. 1999, Lyman et al. 1999, Dujardin et al. 1999). The clear-cut shape divergence disclosed here between the prolixus-robustus group and the stali-pictipes group (Fig. 2) was in accordance with this evolutionary classification, illustrating again that the metric characters might be able to portray the main evolutionary relationships within the genus Rhodnius (Dujardin et al. 1999). To verify the robustness of the identification procedure, we used the same technique as explained before using as supplementary data randomly selected, known individuals of either R. stali, R. pictipes, R. prolixus, or R. robustus. This test was repeated one hundred times, with no one case of wrong re-classification. Thus, this study confirmed the presence of R. robustus in Bolivia, as already suggested by Lent and Jurberg (1969). It also highlighted the relevance of morphometrics to taxonomically interpret one individual when related species data are available for comparison. ACKNOWLEDGEMENTS To Aldo Valente, Michael Miles, and Chris Schofield, for providing us with representatives of different Rhodnius species. To Dr R Andrade, Director of the INLASA (La Paz, Bolivia) and Dr S Mollinedo, Director of the Department of Parasitology, INLASA, for helping this investigation. REFERENCES
This work has benefited from international collaboration
through the ECLAT network. Copyright 2001 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc01184f2.jpg] [oc01184f1.jpg] [oc01184t1.jpg] [oc01184t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}