 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(7) 2001, pp. 965-972 Specific Antibody Levels and Antigenic Recognition of Wistar Rats Inoculated with Distinct Isolates of Trypanosoma evansi Adriana O Queiroz, Ana P Legey, Samanta CC Xavier, Ana M Jansen+ Departamento de Protozoologia, Instituto Oswaldo
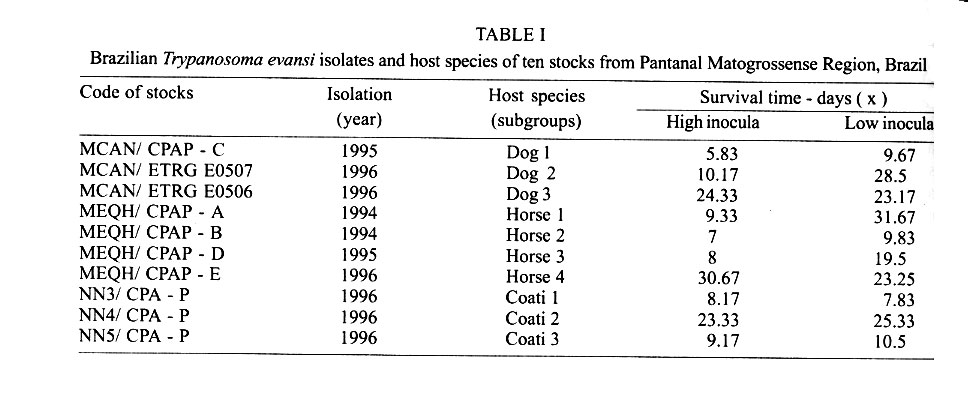
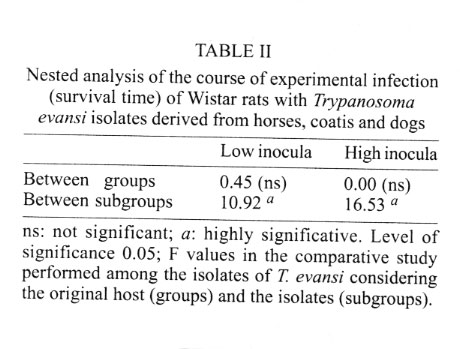
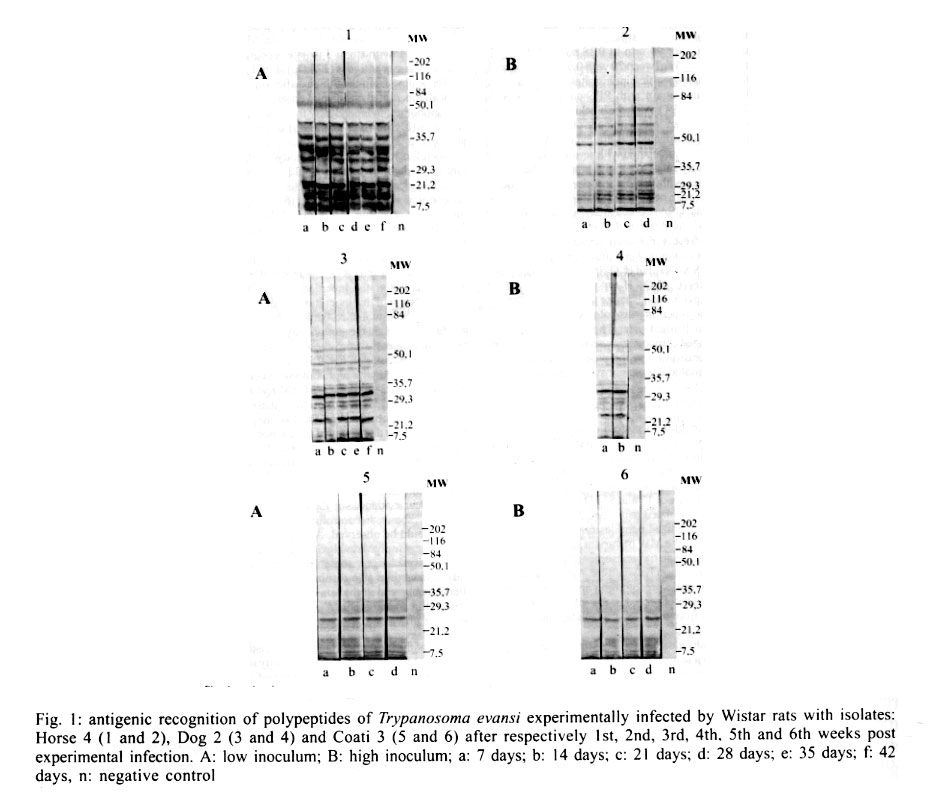
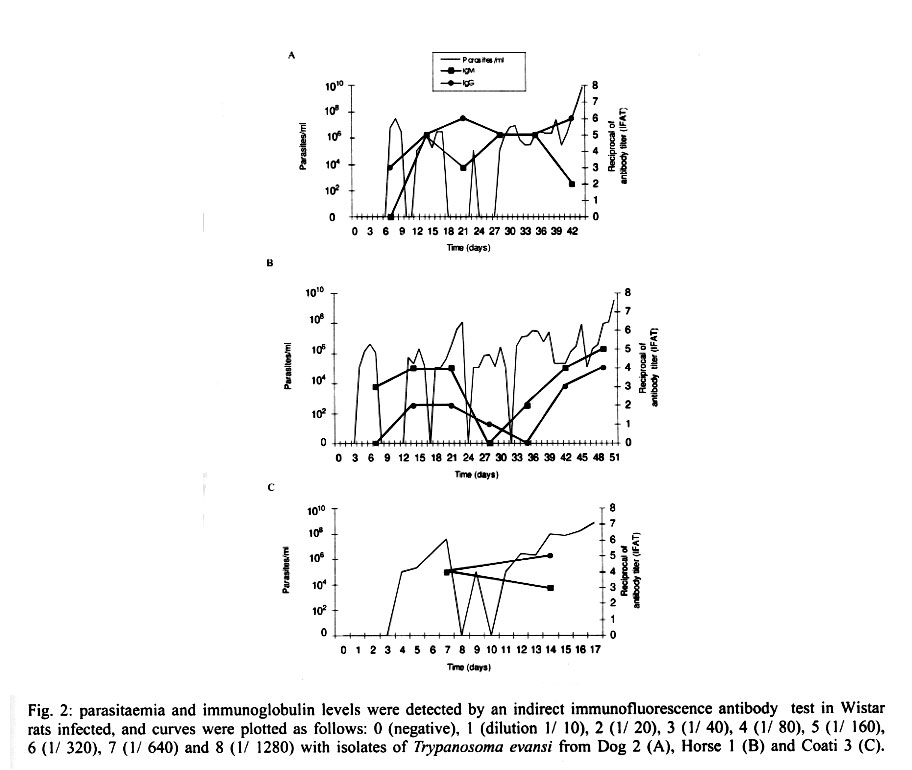
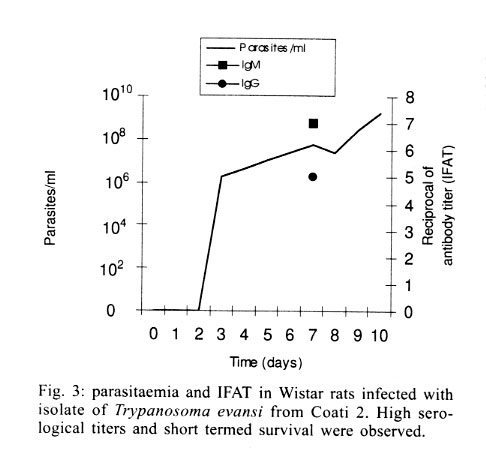
Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil Received 5 September 2000 Code Number: oc01188 "Mal de Cadeiras", an enzootic disease caused by Trypanosoma evansi, is one of the most important trypanosomiases in the Brazilian Pantanal region. The disease affects mainly horses, which are widely used in extensive cattle production, an activity of greatest economical significance for the region. The parasite also infects sylvan (coatis and capybaras) and domestic (dogs) animals, respectively considered wild and domestic reservoirs of T. evansi. For a better understanding of the interaction of T. evansi with its rodent host, we evaluated the differences in the specific antibody level patterns and in the parasitic peptides recognition patterns of experimentally infected Wistar rats. The rats experimentally infected with T. evansi isolates obtained from coatis, dogs and horses were submitted to indirect immunofluorescence test (IgM e IgG) and Western blotting. The serological titers for IgM and IgG ranged between 1:40 and 1:160. The most recognized polypeptide profiles were in a range of 17 and 74 kDa. Our data suggest that the humoral immune response in Wistar rats is not sufficient for granting an effective control of T. evansi infections. Key words: Trypanosoma evansi - Wistar rats - antigenic recognition - indirect fluorescent antibody test Trypanosoma evansi is a widely distributed hemoflagellate, closely related to T. brucei. However, unlike other salivarian trypanosomes, T. evansi is mechanically transmitted by blood sucking insects, especially by Tabanus spp. and Stomoxys spp. In South and Central America, T. evansi is also transmitted by the vampire bat (Desmodus rotundus) which also acts as a carrier (Hoare 1972). Since only the slender forms have been described, T. evansi is considered a monomorphic trypanosome. Up to the present, many T. brucei related species have been described based on differences regarding virulence, morphology and hosts, among others T. equinum, T. venezuelense and T. hippicum in South America and T. marocanum in Morocco. The taxonomic significance of these species is at present considered dubious. Although T. evansi is presenting geographic and epidemiological peculiarities, only one taxon is accepted. This point of view was confirmed through recent molecular and biochemical characterizations (Borst et al. 1987, Stevens et al. 1989, Lun et al. 1992a, b). In spite of the susceptibility of almost all species of domesticated livestock to T. evansi, the main host varies according to the geographical region. In Central and South America, the disease affects mainly horses, the principal hosts in Africa are the camels and the main host in China is the pig (Dia 1997). Similar to T. brucei, T. evansi multiplies during a short period after infection in the inoculation site, creating a local lesion described as a chancre. After one or two weeks, the parasites reach the bloodstream and in more advanced stages of the infection they cross the meningeal barriers (Luckins et al. 1992). Parasitaemia waves have been noticed in experimentally infected animals but only a limited set of variable antigenic types have been described in T. evansi (Jones & McKinnell 1985). The humoral immune response of the host in salivarian trypanosome infections is considered very important, and the recognition pattern of some given parasitic antigens by the host has been related to the control of parasitaemia waves (Shapiro & Murray 1982, Dempsey & Mansfield 1983). Despite its broad distribution and the economical losses it causes, T. evansi is far less studied than the Glossina transmitted trypanosomes. Very few data are available about the transmission dynamics in the natural environment, such as the factors that determine outbreaks of the disease and the course of natural and experimental infections. Consequently, for a better understanding of this enzootic disease we need biochemical, immunological, molecular and biological data of T. evansi derived from distinct hosts and geographical regions. Only limited antigenic diversity has been described among T. evansi isolates. This characteristic reinforces the importance of the host's humoral immune response and suggests the possibility of controlling this enzootic disease through immunoprophylactic methods (Jones & McKinnell 1995). While there are many studies on T. brucei, only few data on T. evansi antigen recognition are available and none, regarding Brazilian isolates (Uche et al. 1992, 1993, Uche & Jones 1994). The importance of antibodies in the control of parasitaemia by T. evansi is poorly known and still controversial (Franke et al. 1994, Aquino et al. 1999). There is only one single study on antigenic variation in Wistar rats. This study, however, was carried out with T. evansi isolates from India and used parasitic flagellar and cell membrane peptides (Singh et al. 1995). In the present study we examined T. evansi antigen recognition and specific antibody levels of Wistar rats (Rattus norvegicus) experimentally infected with domestic and sylvatic T. evansi isolates from the Brazilian Pantanal region. MATERIALS AND METHODS Trypanosomes - T. evansi isolates from domestic and sylvatic animals (Table I) from the Pantanal of Mato Grosso were studied. The coati and dog isolates were obtained by one member of our group (AM Jansen) and horse isolates were kindly supplied by RAMS Silva (Embrapa, MT). The origin and codes of stocks used in the present work are given in the Table I. It is worth mentioning that Horse 4 and Coati 2 isolates are the most and the less virulent T. evansi isolates, respectively (Queiroz et al. 2000a). All isolates were collected in the Southern region of the Pantanal of Mato Grosso. After being collected, the isolates were maintained in liquid nitrogen until use. Preparation of trypanosome antigens - Total antigen was prepared from trypanosomes collected from Wistar rats experimentally infected and previously immune-suppressed (cyclophosphamide, 200 mg/kg body weight). At the point of fulminating infection (109 parasites/ml), the rats for each isolate were exsanguinated and the parasites were purified by separation in a DEAE cellulose column and re-suspended in ice-cold phosphate saline glucose (PSG) pH 8.0 (Lanham & Godfrey 1970). Trypanosomes were disrupted in five to six cycles of 2 min/4°C in an ultrasonic disintegrator (Branson – 2510, USA). The parasites were then lysed through freezing in Methanol and dry ice, alternately. After this, buffer containing protease inhibitors (Trizma base 20 mM, NaCl 40 mM, EDTA 10 mM, Iodoacetamyde 2 mM, 1,10 Phenantroline 1,6 mM and PMSF 1 mM) was added. The total antigen was aliquoted into 100 µl volumes and stored at -20°C. Inoculations - After purification of parasites from previously infected rats through an anion exchanger column (DEAE cellulose), the inocula of each T. evansi isolate were adjusted in a Neubauer chamber so as to yield, 10 and 103 parasites/g of body weight respectively and injected into young, male Wistar rats weighing 100-150 g. Two batches of six animals per inoculation schedule were inoculated intraperitoneally with 10 flagellates/g of body weight (low inoculum) and 103 flagellates/g of body weight (high inoculum). The follow-up included daily fresh blood smear examinations and weekly blood collection. The sera were aliquoted into 50 µl and stored at -20°C. Sera from non-infected rats were used as negative controls. Serological follow-up - An indirect fluorescent antibody test (IFAT), as described by Camargo (1966) was performed to follow-up T. evansi infected Wistar rats (Table II). The antigen, consisting of tripomastigote forms of T. evansi, was adjusted to 40 parasites by microscopic field examination (40x) and stored at -20oC. Positive control serum was obtained from a rat experimentally infected with strain Horse 4. For the indirect immunofluorescene reaction a conjugate of anti-mouse IgG FITC conjugate (Sigma F6257) and anti-mouse IgM FITC conjugate (Sigma F9259) was used. Due to the non-existence of anti-rat IgM we preferred to use anti mouse conjugate (IgM and IgG). The difference in the serological titers as tested by us was not significant. SDS-PAGE - The trypanosome total antigens were solubilized by boiling them for 4 min in an equal volume of 1X Laemmli's sample buffer, loaded into sample wells and separated by SDS-PAGE under reduced conditions in a 10% polyacrylamide gel containing SDS. The concentration of the stacking gel was of 4.5% polyacrylamide (Laemmli 1970). A mixture of calibration proteins with molecular weight ranging from 7.5 to 202 kDa (molecular weight markers, Broad range, BIORAD) was included in a separate well during each run. All gels were 1.5 mm thick and run at a constant current of 100 V for approximately 2 h. Western blotting - After SDS-PAGE, antigens from four isolates (Horse 3, Horse 4, Dog 2, Coati 3) were electrophoretically transferred overnight from the gel to a nitrocellulose membrane (0.22 µm pore size) (Western blotting) at 0.17 A and 4°C following the technique described by Towbin et al. (1979). The section of the nitrocellulose membrane containing the molecular weight markers and the section containing the strips was cut into strips of 5 mm width. The total strips were immediately stained for 10 min with 0.2% Ponceau S (Sigma, USA) in 3% acetic acid and 97% tri-distilled water to verify all transferred trypanosomal proteins and molecular weight markers. After that, the blots were washed for 30 min with tri-distilled water. The strips were then placed for 90 min in PBS solution (pH 7.2) containing 0.5% Tween 20 with 0.5% defatted milk to block non-specific binding sites. The strips were then washed again with PBS containing 0.05% Tween 20 (PBST). After the blocking, the strips were incubated overnight with sera obtained from experimentally infected rats diluted in PBST (1:200). One strip was incubated with serum obtained from an uninfected rat as a negative control. After six 15 min washes with PBST the strips were incubated for 90 min with peroxidase conjugated anti-mouse IgG (Sigma, USA) diluted in a ratio of 1:4000. After six washes as described above, the strips were added to freshly prepared substrate solution (15 mg of 3,3'- diaminobenzidine tetrahydrochloride (Sigma, USA), 40 µl of H2O2 in 60 ml of citrate phosphate buffer (C6H8O7, H2O – 0.01M and Na2HPO4 – 0.02M) pH 5.0 to visualize the antigenic components. The reaction was interrupted by washing the blots in distilled water. The molecular weight of the visualized components was measured by means of a calibration curve prepared for each isolate based on molecular weight standards (Singh et al. 1995). RESULTS The recognition pattern of experimentally T. evansi infected rats is summarized in Fig. 1. Experimentally infected Wistar rats recognized polypeptides of molecular weight ranging between 7 and 200 kDA, but the more intense recognition occurred in a range between 17 and 57 kDA. With exception of the rats inoculated with Coati 3 isolate, all tested animals recognized peptides ranging between 53 and 63 kDa – the MW of variant surface glycoproteins of salivarian trypanosomes (Richards 1984). No correlation between the number and/or intensity of peptide recognition and the serological titers could be observed in the infected rats, independently from the T. evansi isolate or inoculation schedule used. No individual differences in the recognition pattern in a same batch of rats inoculated with the same T. evansi isolate were observed, but it differed in the different rat groups according to the inoculated T. evansi isolate . No variation could be observed during the course of the experimental infection. Actually, the profiles remained unchanged since the first testing (with seven days) (Fig. 1). Rats infected with what we call high or low inocula of Horse 4 survived significantly longer and recognized a greater number of T. evansi antigens (Tables I, II). Moreover, no further differences were observed in the recognition pattern of rats infected with high or low inocula (Fig. 1). Rats infected with the Dog 2 isolate recognized antigens with the molecular weight of 18, 27, 48, 63 kDa and rats infected with Coati 3 isolate recognized significantly lesser antigens (22 kDa) (Fig. 1). Beginning from the 7th day after infection, almost all inoculated rats displayed specific IgM and IgG antibodies, independently from the inoculation schedule (high or low inoculum) or origin of the isolate (Fig. 2). As refers to the specific antibody levels, important individual differences could be observed in the serological titers of the infected rats. However, no correlation between these differences and the inoculation schedule, the origin of the inoculated T. evansi isolate or the recognition pattern could be observed. Most of the inoculated rats presented serological titers for IgM and IgG in the serum already from the 7th day after the inoculation, independently from the origin of the inoculum. No direct correlation could be established between antibody (IgM and IgG) levels and the survival time of the infected rats, independently from the origin of the isolate and/or the inoculation conditions. Rats infected with high and low inocula of the ten T. evansi isolates presented IgM and IgG titers ranging between 1/40-1/160. The highest serological titers were observed on the 14th day after inoculation in rats that received a high inoculum with Dog 3 isolate (IgM: 1/640 and IgG: 1/320) and on the 21st day after inoculation in rats that received a low inoculum with Horse 4 isolate (IgM:1/1280 and IgG: 1/640). No clear evidence for the effectiveness of the immune response in controlling the parasite populations could be observed. Although higher serological titers for IgM and IgG were observed in rats that survived longer, some rats that survived for only a short time also displayed high serological titers as observed in the rats infected with Coati 2 isolate (Fig. 3). DISCUSSION One of the most important characteristics of the salivarian trypanosomes is their capacity to modify their surface coat during the course of infection (Overath et al. 1994). The African trypanosomes variant surface coat (VSG) is reported to be a single 10-15 nm thick coat of glycoproteins with a MW ranging between 53 and 63 kDa (Richards 1984). In our experiments, all animals recognized a peptide in this MW except the rats inoculated with Coati 3 isolate that showed the shortest survival time. Furthermore, it is worth mentioning that rats inoculated with Coati 3 isolate, one of the most virulent isolates, recognized a polypeptide of 21 kDa, whose importance is not mentioned in the literature. Although unlikely since only observed in rats infected with this isolate, it can not be ruled out that the absence of recognition of polypeptides ranging between 53-63 kDa could be the result of an alteration of the parasite surface during the purification procedure. The surface coat of Brazilian T. evansi isolates has never been characterized, but presumably, it is in the same range of MW already described for T. brucei and other T. evansi isolates. T. evansi is an interesting trypanosomatide. Despite its broad geographical distribution, wide range of hosts and distinct epidemiological features it is recognized as homogeneous species (Borst et al. 1987, Stevens et al. 1989). Indeed, genetic diversity as measured by the conventional biochemical and molecular markers has always led to irrelevant results (Masiga & Gibson 1990, Lun et al. 1992b). In a previous work we described the homogeneity of ten T. evansi isolates derived from domestic and sylvatic mammals by means of biochemical (isoenzymatic profiles) and molecular (schizodeme analysis) characterization. In spite of this homogeneity we observed significant differences in the virulence pattern of the T. evansi isolates, which could not be correlated with the species of the original host (Queiroz et al. 2000a, b). In the present paper, we observed differences in the recognition pattern in the different batches of rats inoculated with the distinct T. evansi isolates. These differences are probably due to variations of the polypeptide profile of the parasite isolates and not to individual variations in recognition since animals from the same batch displayed the same recognition pattern. These data show that a correlation of genotypic and phenotypic characteristics of parasites with distinct epidemiological features is still a challenge and indicate that the multiple influences of environmental factors on both, parasites and hosts, should also be taken into account. Differences between resistant and susceptible rabbits with respect to their recognition pattern to non-surface components of T. evansi were observed: the recognition of polypeptides of 61 kDa, 67 kDa and 94 kDa, respectively, by the rabbits was associated to resistance (Uche et al. 1992). In our experiments, no recognition of peptides with this molecular weight was observed, suggesting peculiarities in the interaction of T. evansi with its different host species. In T. congolense and T. brucei infections, a correlation in the recognition pattern between resistant N'Dama and susceptible Boran and Zebu cattle could also be noticed (Authié et al. 1993). This was not the case in this work, since all isolates of T.evansi were highly pathogenic for the Wistar rats and 100% mortality was observed. It is worth mentioning that the rats inoculated with Horse 4 isolate survived longer and recognized more antigens, mainly in a high molecular weight range. Notwithstanding, all animals died with patent parasitaemia (Fig. 1, Table II). The homogeneity in the recognition pattern in the early stages of infection in comparison to the final stages suggests that the fate of the infected animals was determined in the early stages of the infection. It already was described for T. evansi and other salivarian trypanosomatids in other hosts, that the humoral immune response, mainly IgM, plays an important role in the control of the circulating parasites (Roelants & Pinder 1984, Uche & Jones 1994). In our experiments, this correlation was not so clear since the infected rats displayed comparable antibody titers. Our data confirm the findings of Aquino et al. (1999) who observed that high levels of parasitaemia persisted throughout the experiments in spite of the high antibody titers for IgM and IgG. Moreover, in spite of a longer survival, all animals died even when injected with low inocula. This suggests the humoral immune response, at least in this host model, to be far from being sufficient for controlling the parasite populations. It is worth mentioning that the Wistar rats responded earlier and with higher serological titers than previously reported for other animal models such as dogs (Aquino et al. 1999), rabbits (Luckins et al. 1978), camels (Luckins et al. 1979) and guinea pigs (Oliveira et al. 1989). Probably there exist characteristic mechanisms for controlling the parasitaemia in each animal species. Uche and Jones (1994) described that rabbits recognized a smaller spectrum of antigens than rats, reinforcing the idea of characteristic control mechanisms for T. evansi infection of each animal species. These peculiarities could be at least one of the reasons for the distinct aspects of the disease. Indeed, the enzootic disease caused by T. evansi displays distinct features according to the geographical region. Pigs are the main reservoir in Asia, camels in Africa and horses in South America. The Chinese isolates, on the contrary to the others, are cultivable in axenic media (Zweygarth et al. 1991). Distinct virulence patterns were also described (Queiroz et al. 2000a). All these factors should be taken into account in case an immunoprophylaxis program is to be planned. It will also be necessary to evaluate the role played by cross recognition and cross reactivity among T. evansi isolates of a same geographical region. ACKNOWLEDGEMENTS To R Mexas for help with the photographs and A Ivo for technical help. REFERENCES
This investigation was supported by Capes, Faperj and Fiocruz. Copyright 2001 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc01188t1.jpg] [oc01188f3.jpg] [oc01188f1.jpg] [oc01188f2.jpg] [oc01188t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}