 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
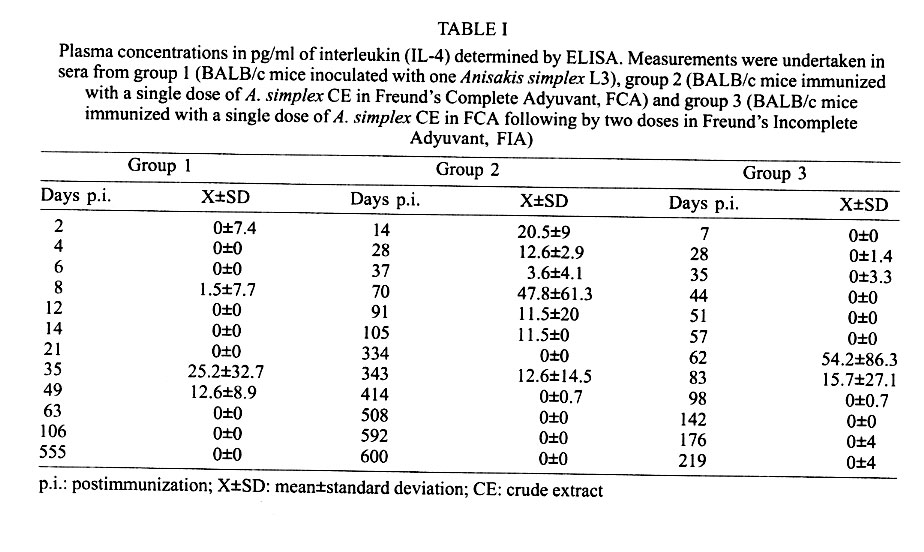
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(7) 2001, pp. 979-982 SHORT COMMUNICATION Interleukin-4 Production in BALB/c Mice Immunized with Anisakis simplex María Jesús Perteguer, Carmen Cuéllar+ Departamento de Parasitología, Facultad
de Farmacia, Universidad Complutense, Ciudad Universitaria, 28040, Madrid, España
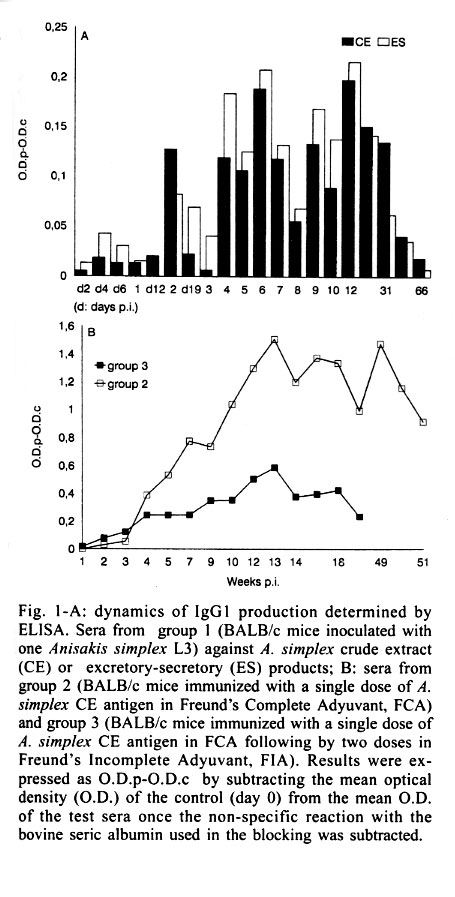
Received 10 October 2000 Code Number: oc01190 We investigated the interleukin (IL-4) levels in BALB/c mice immunized with Anisakis extract in single or multiple doses and in mice orally infected with a larva. From animals immunized maximum responses were obtained with the multiple doses with an only IL-4 peak. Conversely, in the mice inoculated with a larva per os, the IL-4 levels showed two peaks of different rates. Key words: interleukin-4 - BALB/c - Anisakis simplex During the latest years a relation among the different subsets of T cells (Th1/Th2) and the resistence or susceptibility against certain diseases has been almost established. The capability of a host to eradicate with effectiveness any invasive organism depends on the kind of the specific immune response scored (Romagnani 1997). In general terms, the protection against intracellular parasites is carried out by T cell Th1 response, while the protection against extracellular parasites is better leaded by the synergic action of both Th1 and Th2 cytokines together, as it occurs with Th0, for example (Daugelat & Kaufman 1996). Since the specific antibodies showed by the murine BALB/c strain (susceptible strain) indicated a Th2 response after the Anisakis infection (Perteguer & Cuéllar 1998), we proposed the investigation of the interleukin (IL-4) levels (marker of Th2 responses) in mice immunized with A. simplex crude extract (CE) in a single dose or in multiple doses, as well as, in mice orally infected with a living third stage larva (L3). A. simplex L3 were picked up manually from naturally infected blue whiting (Micromesistius poutassou). A. simplex antigens were prepared as previously described (Perteguer & Cuéllar 1998). Briefly, to obtain excretory-secretory (ES) products L3 were cultivated in Minimum Essential Eagle Medium with Earle's salts (ICN Biomedical Inc., Costa Mesa, CA, USA). For the preparation of CE L3 were homogenized, sonicated and delipidized with n-hexane. Thirty BALB/c mice were divided into three equal groups: group 1 (inoculated with one L3 of A. simplex by oral administration with gastric tubing); group 2 (immunized with a single dose of 1 mg of A. simplex CE in Freund's Complete Adyuvant, FCA, by the intramuscular route) and group 3 (immunized with a single dose of 1 mg of A. simplex CE in FCA following by two doses in Freund's Imcomplete Adyuvant, FIA, given weekly during three weeks). As negative controls animals immunized with adjuvant alone were used. Other 20 BALB/c mice were divided into two equal groups: group 2A (immunized with a single dose of FCA by the intramuscular route) and group 3A (immunized with a single dose of FCA following by two doses of FIA given weekly during three weeks, by the same route). Animals from group 2, 2A, 3, and 3A were bled weekly. Animals from group 1 were bled each two days (during the first 21 days) and weekly for the remainder of the experiment. Specific IgG1 levels were measured by ELISA against larval ES or CE antigen using a goat affinity-isolated, horseradish peroxidase-conjugated antibody specific to mouse IgG1 (CALTAG Laboratories, San Francisco, CA, USA; Perteguer & Cuéllar 1998). Serum IL-4 levels were detected using a commercial ELISA system according to the manufacturer's specifications (Mouse IL-4, Cytoscreen Immunoassay kit, BioSource International, Inc., Camarillo, CA, USA). The patterns of production of IL-4 were different among the three groups of animals tested, with higher responses in the immunized animals than that observed in the infected ones. Within the groups immunized with antigen the maximum responses were obtained after the multiple doses of antigen (Table I). In this case, a single peak was observed (weeks 8-14 p.i.) being correspondent with a gradual increase of IgG1 levels detected by ELISA (Fig. 1B). No production of the IgG1 subclass was detected in the control groups (Table II). Conversely, in the mice infected orally with one larva, the IL-4 levels showed two peaks of different intensity. One peak with moderate levels, between the days six and 12 p.i. (Table I). By the day 21 (week 3), a second increase of IL-4 was produced, although very higher than the former. This response was maintained until the week 9 (Table I). On the other hand, other two peaks were seen against the ES antigen (weeks 3-8 p.i., 9-31 p.i.) (Fig. 1A). When the IL-4 production patterns were studied, the higher responses were observed in the immunized animals when compared to those obtained in the infected ones. The maximum responses were obtained within the group immunized with the multiple doses of antigen. This fact was consequent with the higher concentration of antigen used in this experiment. IL-4 peaks were correspondent with gradual increases of IgG1 levels detected by ELISA. In the case of the mice infected orally with one larva, we observed one peak with moderate levels, between the days six and 12 p.i., probably due to the stimulation induced by the ES products released from the living larva. This fact was confirmed by the observation of a correlative increase of IgG1 (IL-4 mediated/Th2-dependent) when the optical density against the ES antigen were measured by ELISA (Fig. 1A). On the other hand, the second increase of IL-4 was observed probably due to the stimulation of the somatic products released from the dead larva. Other two peaks were seen against the ES antigen, probably also due to the somatic antigen released from the dead larva, being, their products, capable of recognizing the ES antigen due to its partial antigenic community. Newly, we seen that the response of IgG1 against A. simplex is mainly due to the products released from the dead larva showing their capability of inducing immediate hypersensibility reactions Traditionaly, it is accepted that in the immunity induced by helminth parasites, are involved eosinophyls, mastocytes and IgE and it is, consequently, reasonable to assume that the immunity is, in this case, dependent of the Th2 subset of T lymphocytes (Finkelman et al. 1991). The IL-4 contributes to the control of the infections, by the induction of several effects, as the stimulation of the IgG1 production, the inhibition of the sinthesis of the inducible nitric oxide sinthasa by the macrophages, etc., although the IL-4 could be only one of the multiple stimuli necessary of the protective immunity against nematodes. Is necessary observed that all the cytokines associate to Th2 responses are not protective to the host. At a distance of the evolution, hosts have developed capabilities to recognize the characterictics of parasitic nematodes as a factor able to arise a serotype response of cytokines, but they have not developed the capability of distinguishing individual characteristics. As consequence, hosts initiate a conjunct of defence mechanisms that controlled the infection in the greater part of members of these community, but they cannot be necessary for the defence against a particular parasite and, inclusively, can cause damage to the host (Finkelman 1995). These "parasitic characteristics" are comparted by numerous allergens, and may be responsible of the poor adaptation developed against molecules, such as, the proteases of the dust acari, and the phospholipids of the bee venom or the pollen antigens (Finkelman 1995). Such, it could be the case of the Anisakis antigens which have been implicated as cause of allergic disorders (Audícana et al. 1995). In a general manner, Th2 responses are protective against Heligmosomoides polygyrus infections, neutral against Nippostrongylus brasiliensis and blokader of the protective responses in Trichinella spiralis infections when Th1 and Th2 responses appeared simultaneously but in different locations (Scott & Kaufmann 1991) and in the Schistosoma mansoni ones (Finkelman et al. 1991). In our experimental model, a Th2 response, as it is induced in the BALB/c strain, does not appeared to be protective, although a predominance of this response close to a Th1 stimulation contribute to the resistence in the C57BL/10 strain (Perteguer & Cuéllar 1998). REFERENCES
Copyright 2001 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc01190t1.jpg] [oc01190t2.jpg] [oc01190f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}