 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
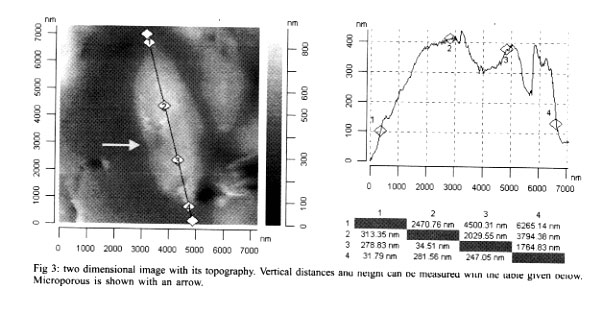
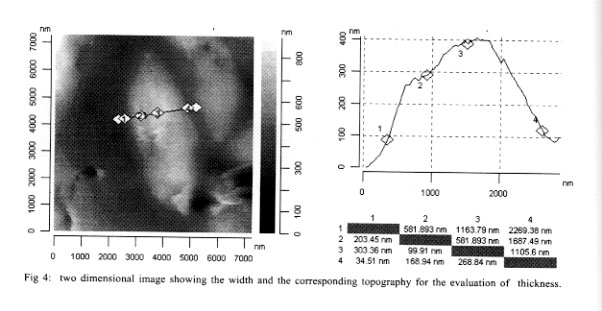
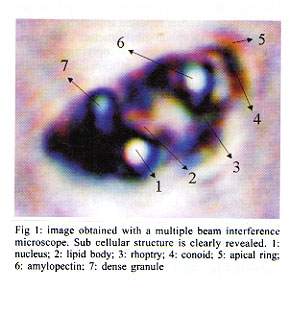
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(7) 2001, pp. 983-986 Morphological Investigation of Toxoplasma gondii in Vivo by a Multiple Beam Interference Microscope H Medina, JM Barboza, H Urdaneta*, M Rondon*, NV Joshi/+ Departamento de Fisiología, Facultad de
Medicina *Instituto de Inmunología Clínica, Universidad de Los
Andes, Merida, Venezuela Received 6 December 2000 Code Number: oc01191 A recently developed technique, namely multiple beam interference microscopy, has been applied to investigate the morphology of the parasite Toxoplasma gondii for the first time. The interference pattern obtained from the multiple internal reflection of a T. gondii, sandwiched between a glass plate and a cover plate, was focused on the objective of a conventional microscope. Because of the enhance contrast, several details of sub cellular structure and separating compartments are clearly visible. Details reveal the presence of a nucleus, lipid body, dense granule, rhoptry and amylopectin. The wall thickness of the membrane of the lipid body and the amylopectin is of the order of 0.02 µm and can be clearly distinguished with the help of the present technique. The same parasite has also been examined with the help of atomic force microscopy, and because of its thick membrane, the inner structural details were not observed at all. Sub cellular details of T. gondii observed with the present technique have been reported earlier only by low amplification transmission electron microscopy and not by any optical microscopic technique. Key words: multiple beam interference microscopy - high resolution - morphology - Toxoplasma gondii - atomic force microscopy A multiple beam interference microscope has been recently developed (Joshi & Medina 2000a) to investigate morphological details and for the examination of internal sub cellular structure of microscopic organisms in living conditions. This technique has been successfully applied to human spermatozoon, Entomoeba histolytica and Candida albicans (Joshi & Medina 2000b). The power of the present technique has been already confirmed when the obtained results were compared with the conventional Differential Interference Contrast Technique, commonly known as DIC (Joshi & Medina 2000b). In the present investigation, we want to extend earlier reported approach to examine in vivo the morphological structure of the parasite T. gondii which causes toxoplasmosis. T. gondii is an obligate intra-cellular parasite and has dimension of the order of 6 x 2 µm and its dimensions are not very much altered from stocks to stocks. Interest has been reemerged in this parasite because it has been found that about 80% of Aids patients die from infections other than human immuno-deficiency virus (HIV) (Kasper & Gatel 1998) and a large fraction of it is due to toxoplasmosis (Lutch & Nord 1996). More over, it is well established that a pregnant woman, having toxoplasmosis is a potential threat for the baby as there are more than 40% chances that baby will get infection. The consideration of the above mentioned aspects and the availability of the new technique encourages the investigation in this direction. The basic requirement for examining the morphology and structural details of a living organism, in addition to a high-resolution microscope, is high contrast, which ultimately improves the resolution. Increasing the contrast is a key issue in these systems, as there is a very soft contrast between different sub cellular components and organelles. Certainly, DIC improves the contrast, but it is not enough to reveal many structural details. This impediment has been recently overcome by using multiple beam interference microscope in which a ray of light is allowed to pass several times through different organelles and because of different thickness and different proteins (i.e., different refractive index), the optical path lengths are altered. This causes a phase shift and hence amplitude differences which lead to optical contrast. It has been found that the technique based on this principle has a remarkable effect on the capacity to reveal the internal structure with high contrast. Earlier work (Joshi & Medina 2000b) on ameba confirms this view. We have, therefore, extended our work to T. gondii. The obtained morphological details were compared with the ultra structure obtained from electron microscopy and also obtained with atomic force microscopy, which was carried out in our laboratory for this purpose. MATERIALS AND METHODS Culture - T. gondii parasites were cultured and maintained in our laboratory by a conventional method. Every 72 h tachyzoites were injected into female NMRI mice with standard procedures. These naive, four weeks old mice were infected by the intraperitoneal route with 105 tachyzoites. The peritoneal parasites were released by lavage with 3 ml of phosphate-buffered saline (PBS) with pH 7.2 and mounted on a standard microscopic glass slide. Multiple beam interference microscopy - As mentioned earlier, we have used a multiple beam interference microscope rather than a conventional confocal microscope as the confocal effects were not the primary importance. Therefore, the system was not operated in the confocal scanning mode. Moreover, the scanning process, even though is slow, does not permit to reveal all the details of the image which can be photographed with a few minutes of exposure. A morphological details at a resolution of microns were examined by using a recently developed multiple beam interference contrast microscopy (Joshi & Medina 2000a). It essentially consists of a multiple beam interferometer connected to a conventional inverted microscope (Olympus IX-70). This system dramatically increases the contrast and, hence, the resolution in x and y direction. The resolution in z direction is enhanced due to the spatial distribution of the interference pattern and it depends upon the wavelength. Internal multiple reflections were achieved by placing a biological sample (T. gondii) with a fluid having a higher refractive index with respect to the air, between a microscopic slide and a cover plate. This creates multiple reflections, which are brought to the focal plane of the objective of the microscope. The intensity of the transmitted radiation is periodic with thickness and depends directly on airy function. Therefore, the thickness and reflection coefficient were adjusted for certain morphological details. It is worth mentioning that the use of a dye or staining is avoided with the present technique. The other experimental details of the multiple beam interference microscope are given in our earlier publications (Joshi & Medina 2000b). Atomic force microscopy - For ultra structural study, we have employed an atomic force microscopic unit attached to a conventional Olympus microscope (BX 60). The system is commercially known as SIS - ultraobjective obtained from Surface Imaging System. We have used silicon cantilevers of 225 µm length and 30 µm width. The spring constant of the cantilever was 0.001 Nm-1. The outer membrane of T. gondii is very thick and hence, non contact mode was found to be inappropriate. The system was, therefore, operated in contact mode. The unit was installed on a vibration free table and the complete system guaranteed reproducibility. Ultra structural studies of T. gondii, obtained by the earlier mentioned method, was used for the present investigation. The obtained images are given in Figs 2, 3 and 4. RESULTS AND DISCUSSION Fig. 1 shows the images obtained with a multiple beam interference microscope. Several sub cellular structures like nucleus, amylopectin, lipid body, rhoptry, and dense granule can be clearly observed which are not possible to detect by a conventional or DIC microscopic techniques. A complete morphological investigation has been carried out earlier by using low amplification transmission electron microscopy (Dubey et al. 1998) and the results presented here are comparable with it in majority of cases. In some cases, because of low amplification, assignment is tentative. Amylopectin and the lipid body have dimensions of the order of 0.4 µm and they have been unambiguously detected. The most important aspect of this image is that the thickness of the cell membrane of amylopectin is clearly distinguished. In fact, the thickness is of the order 0.02 µm, much less than the conventional Rayleigh detection limit. The high resolution is achieved because of high contrast and we believe that it is a consequence of the present technique. However, the conoid of tachyzoite was not detected even though its length is generally of the order of 0.3 µm. This may be due to the fact that the proteins involved in the apical ring and conoid of tachyzoite have approximately the same refractive index and hence the same optical path; which makes a low contrast between them. In the present investigation we have used the property of different refractive index of the proteins and hence the different path lengths to separate them. No attempt has been made to separate surface and bulk proteins, and identify their structure and function. Such work has been already reported (Tomavo 1996). In order to examine surface topography and if possible, inner structure of T. gondii, atomic force microscopic investigation was carried out and the results are shown in Figs 2, 3 and 4. Fig. 2 shows the three dimensional image of T. gondii. Even though, some structure is seen roughly, it is not possible to identify any of them. In the present case, sub cellular structure, even the gross structure like nucleus or rhoptry, was not detected and this might be due to the thickness of the wall membrane. Several attempts were carried out to see more details but they were not successful. However, the figure is useful to measure the dimensions of T. gondii with precision. Figs 3 and 4 show two-dimensional images recorded with the same experimental method. The length of T. gondii is found to be 6,265 nm (see row 1 and column 4). Here the apical ring and micro porous (shown by the arrow) can be seen clearly. The maximum height is of the order of only 400 nm and the topography shows unevenness because of the sub cellular structure which is located very close to the membrane. As expected, the rate of increase in height is not symmetrical because of uneven distribution of organelle. Fig. 4 shows the width and is of the order of 2,269 nm at the center (see row 1 column 4). Here also the morphology shows the presence of some sub cellular structure, however, it is not possible to identify because they are situated deep inside. Comparison of Fig. 1 with Figs 2, 3 and 4 clearly shows that multiple beam interference microscopy seems to be an useful technique, for examining sub cellular structure, particularly when the cellular wall is very thick. The images obtained with the multiple beam interference microscope can be compared with those obtained with transmission electron micrograph at low amplification. Results show with clarity different structures where optical path difference (thickness or the proteins with different optical index) is altered. In Fig. 1 the structure of the nucleus, lipid body, dense granule, rhoptry and amylopectin can be seen. However, structures like electron dense granule are not detected because the contrast achieved is not enough. The present investigation also shows that ultrastructural study of a parasite with thick membrane with atomic force microscope, provides data about the thickness, the overall dimensions with the precision of a few nm and the most important details of topography. Our present approach had helped us to view sub cellular components in a living condition. Such study, when extended to motility aspect, will turn out to be unique for this purposes. REFERENCES
This work received financial support from CONICIT (project no. G-97000820) and CDCHT (project no. M-681-00-03-B) of University of Los Andes, Merida, Venezuela. Copyright 2001 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc01191f1.jpg] [oc01191f3.jpg] [oc01191f4.jpg] [oc01191f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}