 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(7) 2001, pp. 1005-1011 Variation in Susceptibility to Benznidazole in Isolates Derived from Trypanosoma cruzi Parental Strains VM Veloso, CM Carneiro*, MJO Toledo, M Lana*, E Chiari**, WL Tafuri,MT Bahia/+ Departamento de Ciências Biológicas,
ICEB *Departamento de Análises Clínicas, Escola de Farmácia,
Universidade Federal de Ouro Preto, Campus Universitário, Morro do Cruzeiro,
35400-000 Ouro Preto, MG, Brasil **Departamento de Parasitologia, ICB, Universidade
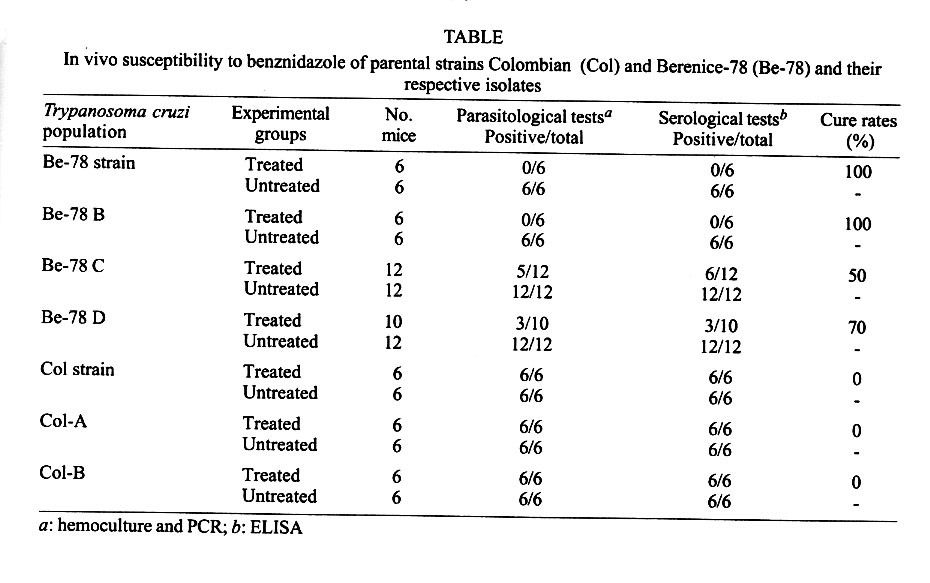
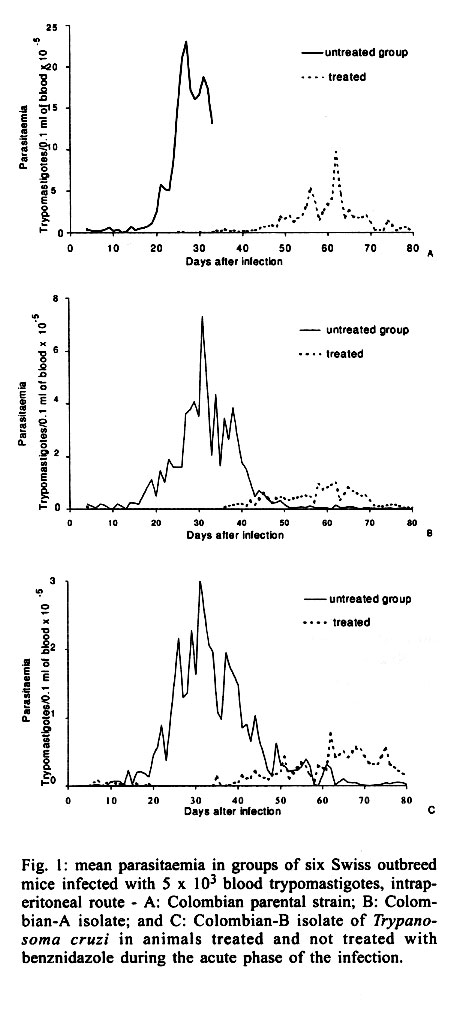
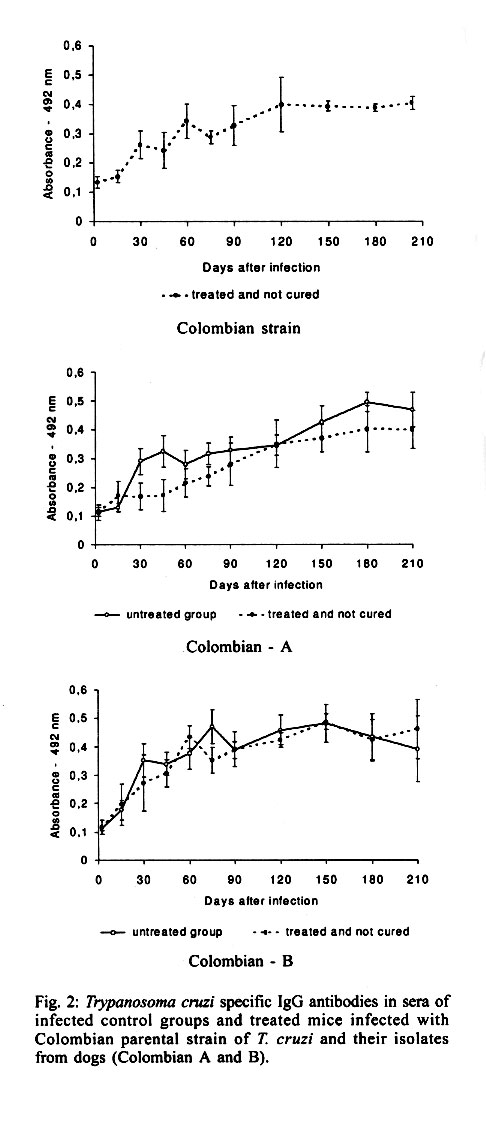
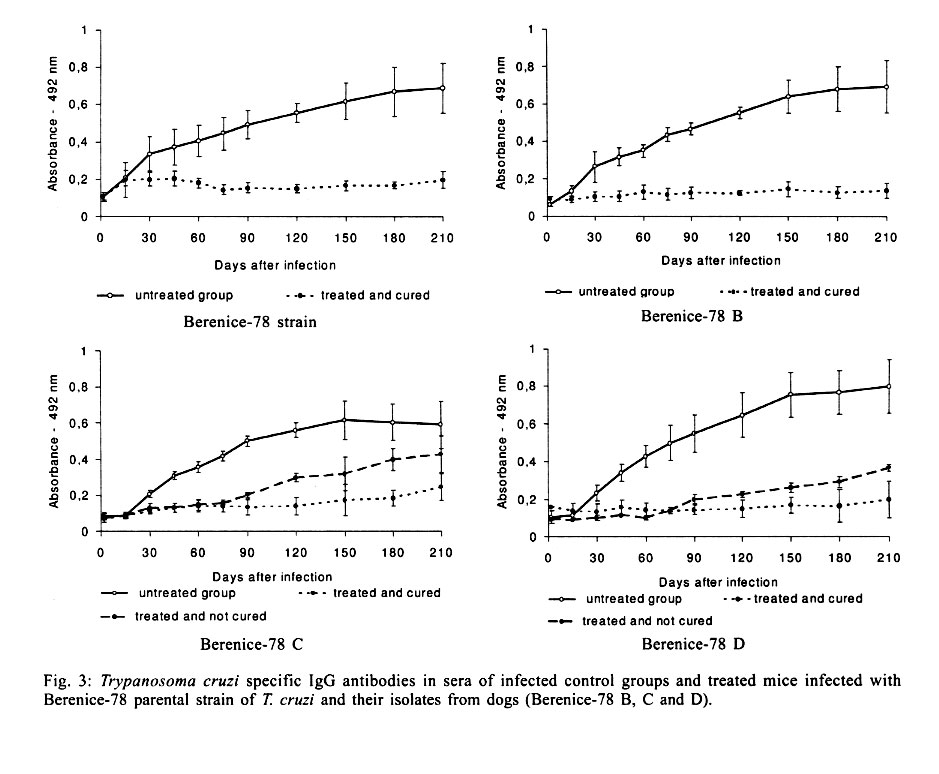
Federal de Minas Gerais, MG, Brasil Received 14 November 2000 Code Number: oc01195 In this work, the susceptibility to benznidazole of two parental Trypanosoma cruzi strains, Colombian and Berenice-78, was compared to isolates obtained from dogs infected with these strains for several years. In order to evaluate the susceptibility to benznidazole two groups of mice were infected with one of five distinct populations isolated from dogs as well as the two parental strains of T. cruzi. The first group was treated with benznidazole during the acute phase and the second remained untreated controls. The animals were considered cured when parasitological and serological tests remained persistently negative. Mice infected with the Colombian strain and its isolates Colombian (A and B) did not cure after treatment. On the other hand, all animals infected with Berenice-78 were cured by benznidazole treatment. However, 100%, 50% and 70% of cure rates were observed in animals infected with the isolates Berenice-78 B, C and D, respectively. No significant differences were observed in serological profile of infected control groups, with all animals presenting high antibody levels. However, the ELISA test showed differences in serological patterns between mice inoculated with the different T. cruzi isolates and treated with benznidazole. This variability was dependent on the T. cruzi population used and seemed to be associated with the level of resistance to benznidazole. Key words: chemotherapy - drug susceptibility - Trypanosoma cruzi - benznidazole Trypanosoma cruzi, the agent of Chagas disease, exhibits considerable biological variability (Carneiro et al. 1991, Andrade & Magalhães 1997). Studies using cloned or uncloned populations, reinforce the heterogeneity of the parasite and demonstrate that, in general, strains of T. cruzi are composed of subpopulations with distinct characteristics (Postan et al. 1986, Finley & Dvorak 1987). Several authors have extensively investigated the correlation between T. cruzi genetic background and the different clinical forms of Chagas disease, as well as biological characteristics, such as virulence, pathogenicity and susceptibility to drugs (Revollo et al. 1998, Andrade 1999). Filardi and Brener (1987) and Toledo et al. (1997) described the existence of strains naturally resistant and non-resistant to benznidazole and ni-furtimox. Natural resistance to nitro-derivates has been suggested as an important factor in explaining the low rates of cure detected in treated patients (Murta & Romanha 1998). Several authors have shown that resistance of T. cruzi strains to benznidazole and nifurtimox increased when parasites were isolated from mice previously treated with these same drugs (Marretto & Andrade 1994, Murta & Romanha 1998). The authors suggest that the initial treatment eliminated the sensitive parasites, preserving the resistant ones, which multiplied and dominated the population. In this manner, the imposition of natural and artificial pressure can result in the selection of a subset of the population. If the strains of T. cruzi present clonal structure, based on the existence of clonal lines without sexual interactions (Tibayrenc & Ayala 1988), it is possible to think that clones with variable degrees of drug susceptibility presented among the whole circulating population would show different growth rates in the host. Consequently, genetic characteristics of the host could favor the development of certain clones within the host, which could reflect differences in the susceptibility to drugs. To confirm this hypothesis, the susceptibility to benznidazole of different isolates of T. cruzi, originated from dogs infected 2, 7, 8 and 17 years before, were comparatively studied in mice with the parental Colombian and Berenice-78 strains. MATERIALS AND METHODS T. cruzi strains and populations isolated from dogs - The parental strains Colombian (Col) - T. cruzi I (Federici et al. 1964), resistant to benznidazole (Filardi & Brener 1987) and Berenice-78 (Be-78) - T. cruzi II (Lana & Chiari 1986), 100% susceptible to benznidazole (Toledo et al. 1995), were used as references. T. cruzi stocks were isolated from different dogs after 8 and 17 years of infection with Col strain (Col A and B, respectively) and with Be-78 after 7 (Be-78 B and C) and 2 (Be-78 D) years of infection. Experimental conditions Mice infection - For each strain or stock of T. cruzi, 12 Swiss outbreed mice (30 days old, 18-20 g weight) were infected by intraperitoneal route with 5 x 103 blood trypomastigotes, obtained from infected mice. Six mice were submitted to treatment with benznidazole (N-benzyl-2-nitro-1-imidazo-lacetamide - Roche Company), and six remained as untreated infected controls. The experiments were repeated when the percentage of cure was different between the parental strains and their isolates. Treatment schedules - Animals were treated after the detection of patent parasitaemia, approximately four days post-inoculation. Benznidazole was administered in a suspension made with 4% arabic gum (Sigma) by oral doses of 100 mg of benznidazole/kg, for 20 consecutive days. Animals were considered cured when both parasitological (fresh blood examination, hemoculture and PCR) and serological (ELISA) tests were negative. Parameters evaluated Parasitaemia and mortality - The parasitaemia was followed from the 4th day of infection until the negativation of the parasitaemia by fresh blood collected from the mouse's tail and the number of parasites was estimated as described by Brener (1962). Curves were plotted using the mean of the parasitaemia from six mice. Mortality rate was expressed as a percentage of accumulated deaths within the period of 180 days after the inoculation. Hemoculture - Mice were bled from the orbital venous sinus 30 and 180 days after the end of treatment. The blood was split into two tubes containing 5 ml of Liver Infusion Tryptose medium (Filardi & Brener 1987). The tubes were incubated at 28°C for 30-90 days and microscopically examinated for the presence of parasites 30, 60 and 90 days later. PCR assay - PCR were performed only in samples from animals with negative hemoculture. Mice were bled from the orbital venous sinus and 200 µl of blood were collected. The sample were immediately mixed with 400 µl of 6 M guanidine HCL/0.2 M EDTA solution (Ávila et al. 1991) and stored at room temperature for one week. This mixture was boiled for 7 min to break the minicircles (Britto et al. 1993). DNA extraction was performed according to Wincker et al. (1994), but using 40 µg of Glycogen (Boeringer Mannheim) to precipitate the DNA. The pellets obtained were resuspended in 40 µl of distilled sterile water and stored at 60°C for 1 h. PCR conditions were described by Gomes et al. (1998). The PCR mixture contained 2 µl of DNA solution, 10 mM Tris-HCl (pH 9.0), 75 mM KCl, 3.5 mM MgCl2, 0.1% Triton X-100, 0.2 mM each dATP, dCTP, dGTP and dttp (Sigma Co. Ltd.), 20 pmol of S35(5'AAATAATGTACGGG(T/G) GAGATGCATGA3') and S36(5'GGGTTCGATT GGGGTTGGTGT3') primers (Ávila et al. 1990), 1.0 unit of Taq DNA polimerase (GIBCO), and 40 µl with water. The reaction mixtures were overlaid with 30 µl of mineral oil and subjected to 35 cycles of amplification in a Research Programmable Thermal Controller (MinCycler TM). The temperature profile were 95°C for 1 min for denaturation with a longer initial time of 5 min at 95°C, 65°C for 1 min for primer annealing, and 72°C for 10 min to extend the annealed primers. Five microliters of PCR products were analyzed by electrophoresis on a 6% polyacrylamide gel and visualized by staining with silver salts (Santos et al. 1993). Serological profile - Ten microliters of blood were collected from mice tails every two weeks for three months and monthly thereafter until seven months post-infection in 90 µl de PBS, pH 7.2. The sera obtained were stored at -20°C, and samples from treated and untreated mice were tested in parallel. ELISA test were performed according to Voller et al. (1976). Peroxidase conjugated goat anti-mouse IgG (Sigma) was used. The cut-off was determined by using the mean absorbance from ten uninfected animals plus two standard deviations. RESULTS Parasitological cure - The Table shows the percentage of cure induced by benznidazole after long-term treatment of different groups of mice infected with each parental strains and their isolates from dogs. Animals infected with Col strain and isolates Col A and Col B did not respond to treatment with benznidazole. On other hand, mice infected with Be-78 strain showed 100% of cure after treatment, whereas mice infected with their isolates Be-78B, C and D) displayed different spectra of susceptibility to benznidazole, with 100%, 50% and 70% of cure rates, respectively. The evaluation of the presence of specific antibodies in the sera of these animals after treatment correlated with the parasitological findings. The effect of benznidazole on the reduction of parasitaemia - Mice infected with Col strain and Col-A isolated showed suppression of parasitaemia during treatment, whereas in animals infected with Col-B isolate this parameter was reduced but not completely suppressed. All mice inoculated with Col strain and Col (A and B) isolates showed patent parasitaemia on the 7th day post treatment. Treated animals showed a decrease in parasitaemia levels of 2.5 and 5 times the levels of untreated control (Fig. 1). All mice infected with Be-78 strain and Be-78 (B, C and D) showed a suppression of parasitaemia on the first day of treatment onset, which persisted throughout and after the treatment period. The mortality rate - At 180 days post treatment the mortality rate was 100% in the control group infected with Col strain and 40% in the treated group. There was no mortality in treated and untreated mice infected with Col (A and B) isolates and Be-78 strain as well as their isolates (Be-78 B, C and D). Serological profile - T. cruzi specific IgG antibodies were detected 20-30 days after infection in sera of infected control groups and all animals displayed high levels of antibodies regardless of the strain or isolate used for inoculation (Figs 2, 3). However differences in the serological pattern among mice infected with different populations of T. cruzi after treatment was detected. In all animals infected with Col strain, IgG antibodies were detected from one to 10 days post treatment. In animals infected with Col-A the antibody levels were under the cut-off values (Abs = 0,233) between 30 and 60 days of infection, raising to control levels thereafter. Antibody levels in treated and untreated animals infected with Col B were similar throughout the infection period (Fig. 2). Similar serological profiles were observed in mice infected with Be-78, and isolates Be-78 that responded to treatment with antibodies levels always under the cut-off value. In all animals infected with Be-78 (C and D), that remained infected post treatment, the antibody levels remained under the cut-off (0,233) until 90 and 120 days after treatment slowly raising afterwards but without reaching the same level of untreated controls (Fig. 3). DISCUSSION Natural resistance of T. cruzi to nitro derivates has been described as an important factor to explain the low rates of cure detected in treated chagasic patients (Filardi & Brener 1987). The authors described the existence of strains that are naturally resistant and non-resistant to benz-nidazole and nifurtimox. Relatively few reports have been published concerning the in vivo induction of drug resistance to T. cruzi. Andrade et al. (1977) showed that the resistance to drugs benznidazole and nifurtimox increased when the parasites were isolated from mice previously treated with the same drugs. Regarding this aspect, it is important to mention that the isolates used in this work were obtained from dogs that were not treated, eliminating, in this way, the possibility of influence of the drug on induction of parasite resistance to treatment. In this study we observed stability of the resistance to benznidazole in the populations from the Colombian strain isolated of dogs 8 and 17 years after infection. However, the fact of the animals infected with the Col-B isolate presented patent parasitaemia during treatment suggests the presence of a higher proportion of resistant subpopulation to benznidazole in this isolate in relation to the parental strain and Col-A. This supposition is corroborated by analysis of the T. cruzi specific IgG profile observed in these mice in relation to the groups inoculated with Col strain and Col-A. The anti-T. cruzi antibody production was delayed in mice infected with the parental Col and Col-A while, in mice infected with Col-B, IgG antibodies were detected also during treatment, similar to observed in control untreated groups. Camandaroba (1999) demonstrated the high resistance of Col strain to benznidazole and verified also similar resistance in seven clones of this strain. Andrade et al. (1977) suggested the selection of resistant parasites to explain the parasite persistence in vertebrate hosts after prolonged treatment. To test this hypothesis Murta and Romanha (1998) investigated the development of drug-resistance of T. cruzi Y strain isolated from mice not cured after treatment with benznidazole or nifurtimox. The authors demonstrate the in vivo selection of a population and clones of T. cruzi resistant to benznidazole from the Y strain (50% resistant to benznidazole) and suggest that the resistance of the strain seems to be related to the sensitive/resistant clone ratio in the population. The present work, however, shows a different situation since we observed a change of susceptibility to benznidazole in isolates of the Be-78 strain considered 100% susceptible (Toledo et al. 1995). Mice inoculated with the isolates (Be-78 C and D) from dogs infected for seven and two years with Be-78 showed susceptibility of 50 and 70%, respectively. These results are difficult to be explained based on the selection of a subset of the population. If within the parental population a clone resistant to benznidazole was to be present, the parental Be-78 strain should present some degree of resistance to treatment. Moreover, correlations between the sensibility to benznidazole and nifurtimox and the genetic distance between T. cruzi stocks have been described in vitro (Revollo et al. 1988) and in vivo (Toledo et al. 2000). Also, Andrade et al. (1992) revealed 82% of coincidence between treatment results in patients and mice infected with same strains. Considering that the genetic characteristic may be one of the factors that modulate the benznidazole resistance/sensibility process, the variation of benznidazole susceptibility observed, specially with Be-78 C, may reflect the genetic plasticity observed in T. cruzi demonstrated by several authors. Pacheco and Brito (1999) studying parasite population in mice revealed evidence that polymorphism in minicircles sequences can emerge during infection with a single clone of T. cruzi. MacDaniel and Dvorak (1993) also showed that under stress conditions, or selective pressures, chromossomes and minicircles shown evidences of genetic plasticity in T. cruzi and Leishmania. The authors postulate that errors in DNA synthesis or replication may occur. These errors could be lethal or produce organisms with either reduced or enhanced survival potential depending upon the influence of external modulating variables such as the environment. In Plasmodium falciparum the chromosome size polymorphism among different strains is caused by the acquisition and deletion of mainly repetitive elements in a subtelomeric position (Lanzer et al. 1994). An alternative hypothesis to explain the variability in drug resistance in Be-78 C and D isolates is that drug-resistant mutants arise for genetic exchange. Two studies of multiple T. cruzi isolates from single localities have found some evidence of genetic exchange (Bogliolo et al. 1996, Carrasco et al. 1996). Although clonal propagation may predominate in transmission cycles involving humans, genetic exchange in natural populations might emerge T. cruzi strains with new resultant biological properties, such as virulence and drug resistance. In this study we also demonstrated variability of specific antibody production profile in mice infected with T. cruzi populations with different patterns of resistance to benznidazole. The variation of antibody levels observed between mice treated and not cured, apparently related to the parasite resistance to benznidazole, seems to correlate with the decrease of parasitaemia. According to Marretto and Andrade (1994) initial treatment would eliminate the sensitive parasites, leading to a predomination of a resistant population. Our data suggest that the serological profile is related to the sensitive/resistant clone ratio in the population. In mice infected with a T. cruzi resistant strain, the suppression of the parasitaemia after treatment is slower and consequently the antigenic stimulation of antibody production is higher. In this work we demonstrate the alteration in the benznidazole resistance/sensibility characteristic in the populations isolates of dogs infected with Be-78 strains. However, the biochemical mechanism that underlines drug resistance in isolates of this parasite remains to be elucidated. ACKNOWLEDGEMENTS To Dr Luiz Carlos Crocco Afonso for his painstaking care in English corrections. REFERENCES
This investigation received financial support from Fapemig, CNPq and Ufop. Copyright 2001 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc01195t1.jpg] [oc01195f3.jpg] [oc01195f2.jpg] [oc01195f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}