 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(8) 2001, pp. 1049-1054 Metazoan Parasite Fauna of the Bigeye Flounder, Hippoglossina macrops, from Northern Chile. Influence of Host Age and Sex M Teresa González/+, Enzo Acuña, Marcelo E Oliva* Departamento de Biología Marina, Facultad
de Ciencias del Mar, Universidad Católica del Norte, Casilla 117, Coquimbo,
Chile *Instituto de Investigaciones Oceanológicas, Universidad de Antofagasta,
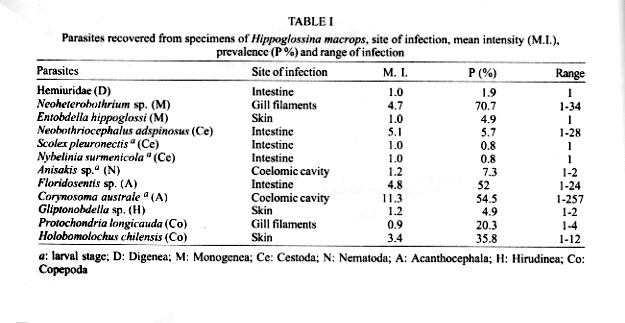
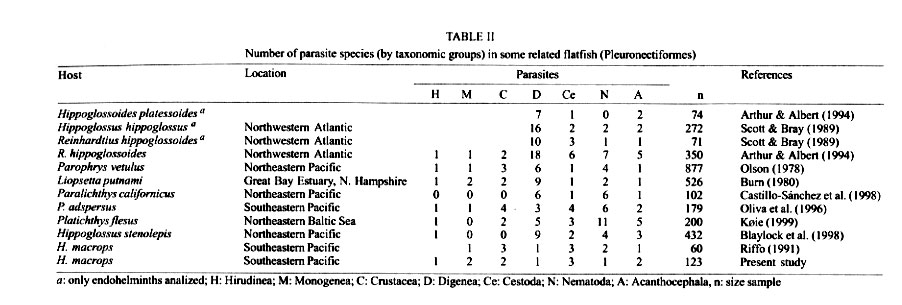
Antofagasta, Chile Received 14 November 2000 Code Number: oc01203 The metazoan parasite fauna of Hippoglossina macrops (n = 123) from northern Chile (30°S) is quantitatively described for the first time, and the role of host age and sex was evaluated. Twelve parasite species were recovered, including 5 ectoparasites (2 Monogenea, 2 Copepoda and 1 Piscicolidae) and 7 endoparasites (1 Digenea, 3 Cestoda, 2 Acanthocephala, and 1 Nematoda). The copepod Holobomolochus chilensis, the monogenean Neoheterobothrium sp., the adult acanthocephalan Floridosentis sp. and the hirudinean, Gliptonobdella sp. are new geographical and host records. The most prevalent ectoparasitic species were the monogenean, Neoheterobothrium sp. and the copepod, H. chilensis. Among endoparasites, the acanthocephalans Floridosentis sp. and Corynosoma australe were most prevalent and abundant. Prevalence and mean intensity of infection for most parasitic species were not affected by host sex, however the prevalence of Floridosentis sp. was significantly greater in males. Intensity of infection was positively correlated with host age for Neoheterobothrium sp., and negatively correlated for Floridosentis sp. and H. chilensis. The helminth species richness of the host H. macrops was lower compared to related flatfishes from the Northern Hemisphere. The relationship of the helminth fauna of H. macrops, its feeding habits and ecological habitats are discussed. Key words: parasites - Hippoglossina macrops - flatfish - ecology - Chile Knowledge about the parasite fauna of marine fishes from the Chilean coast is still scarce, specially for deep water fishes. There are only two quantitative parasitological studies on flatfishes from Chilean coast. Riffo (1991) analyzed the metazoan parasites of Hippoglossina macrops (Steindachner, 1876) off central Chile (36°S), and Oliva et al. (1996) studied the parasite fauna of Paralichthys adspersus (Steindachner, 1867) from northern Chile (23°S). H. macrops is a flounder with a geographical distribution from Mazatlán (Mexico) to Puerto Montt (Chile) (Fischer et al. 1995), occurring from shallow waters to 600 m depth (Yáñez & Barbieri 1974). This fish is an economic resource for local fishermen and it is also a common component of the discard of three important crustacean fisheries, the deep-sea shrimp Heterocarpus reedi Bahamonde 1955, the yellow squat lobster Cervimunida johni Porter, 1903 and the red squat lobster Pleuroncodes monodon (Milne Edwards 1837). Because of the importance of H. macrops, previous studies have been carried out on several aspects of its biology, such as growth (Miranda 1959), reproduction (Voigth & Balbontín 1981) and feeding (Arancibia & Meléndez 1987, Villarroel & Acuña 1999).There are no studies of the parasite fauna of this fish species from northern Chile. The purpose of this paper is to report for the first time the metazoan parasite fauna of the bigeye flounder, H. macrops from the ocean off northern Chile (30°S), to describe parasitism of the host both qualitatively and quantitatively, and to evaluate parasitism with respect to host age and sex. MATERIALS AND METHODS A total of 123 specimens of H. macrops were caught in April 1997 and from March to June 1998. All fishes were obtained from by-catches in the shrimp and squat lobster fisheries. Trawls were done in front of Coquimbo area, between 29°18'S, 71°37'W and 30°50'S, 71°45'W, at depths from 160 to 342 m. Fishes were collected onboard, frozen (-18°C) and transported to the laboratory. After defrosting, fishes were measured (total length, nearest mm), dissected, sexed, and examined for ectoparasites and endoparasites. The parasites obtained were separated and transferred to 10% formalin and sorted and counted by species. Parasite location in the host was recorded. Because the taxonomy of Anisakids is confused, (A. simplex seems to be a species complex, Mattiucci et al. 1997) these were identified to the generic level. Fish age was assigned using the parameters of the Von Bertalanffy growth function. We took into account that growth parameters are different for male and female (males L¥ = 37.29, to = - 0.373, k = 0.325: females L¥ = 41.57, to = - 0.4157, k = 0.281) (Andrade 1999). To analyze if mean age and mean size of male and female fish differed significantly a test-t was used. The influence of sex on prevalence was tested with the "G" likelihood test and the possible effect of sex on mean intensity of infection (previous log n+1, transformation of intensity), was evaluated with an ANOVA and later a HSD Tukey when the ANOVA indicated significant difference. Pearson correlation coefficient (r) was calculated to evaluate the strength of the relationship between host age and intensity of infection. Changes in prevalence of infection with age were tested with the "G" likelihood test. Relationships between mean host size and prevalence were explored with the Spearman's correlation coefficient (rs). Ecological descriptors follow Bush et al. (1997) and the statistical analyses were done according to Zar (1996). Only sexually determined fishes were included in the quantitative analyses, of those 34 were male and 72 were female. Fish ranged from 1.6 to 5.4 years old (x = 2.9 year, SD = 0.83) and between 18 and 33.6 cm (x = 24.5 cm, SD = 3.6). No significant difference between the age of males and females was determined (t = 1.101, df = 102, P = 0.273), but mean size between sexes differed significantly (t = 2.137, df = 102, P = 0.035), the females (25 cm) were larger than males (23.2 cm) RESULTS Twelve parasite species were found in the host H. macrops, including five ectoparasites: Neo-heterobothrium sp. Price, 1943, Entobdella hippoglossi (Müller, 1776) Protochondria longicauda Ho, 1970, Holobomolochus chilensis Cressey & Cressey, 1985 and Gliptonobdella sp. (Sawyer & White, 1969); and 7 endoparasites: Floridosentis sp. Ward, 1953, Corynosoma australe Johnston, 1937, Neobothriocephalus adspinosus Mateo & Bullock, 1966, Nybelinia surmenicola (Okada, 1929), Scolex pleuronectis Müller, 1788, Anisakis sp. Dujardin, 1845 and an unidentified Hemiuridae (Digenea). Site of infection in the host, developmental stage, mean intensity and prevalence are summarized in Table I. The copepod H. chilensis, the monogenean Neoheterobothrium sp., the adult acanthocephalan Floridosentis sp. and Piscicolidae Gliptonobdella sp. are new geographical and host records. The most prevalent and abundant ectoparasite species were the monogenean Neoheterobothrium sp. and the copepod H. chilensis. Among endoparasites, the most prevalent and abundant species were the acanthocephalans, Floridosentis sp. and C. australe (Table I). Ninety three percent of the fish were parasitized with a range of one to 266 individuals, and the species range was one to 6. Eighty-two percent of the infracommunities of H. macrops harbored between two and four parasite species (x = 2.6, SD = 1.1). There was no significant difference in mean intensity and prevalence of parasite species between male and female fish, except for Floridosentis sp., which was significantly greater in males (G = 7.0, P = 0.01). The intensity of infection was positive and significantly correlated with host age for the ectoparasite Neoheterobothrium sp. (r = 0.326, p = 0.005, n = 71). The copepod H. chilensis (r = -0.399, p = 0.017, n = 35), and the adult acanthocephalan Floridosentis sp. (r = -0.405, p << 0.001, n = 52) were significantly but negatively correlated with host age. The intensities of infection for remainding species were not correlated with host age. Prevalence of infection increased with host age for Neoheterobothrium sp. (r = 0.991, P = 0.009, n = 4) and H. chilensis (r = 0.983, P = 0.017, n = 4) while it decreased for Floridosentis sp. (r = - 0.979, P = 0.021, n = 4). Species richness (r = 0.159, P = 0.122, n = 96), ectoparasites (r = 0.205, P = 0.072, n = 78) and endoparasites (r = 0.061; P = 0.604, n = 75) were not correlated with host age. DISCUSSION Rhode (1992, 1993) considered that there is an important effect of latitudinal gradient on metazoan species richness and prevalence of infection of each species, suggesting that it increase towards low latitudes close to the equator. On the other hand, Scott (1982) indicated that parasite species richness is higher in hosts with extensive latitudinal distributions. Riffo (1991) found 11 parasite species in H. macrops from the central-southern Chilean coast (36ºS), while in this study we found 12 parasite species, only 7 of them (58%) previously recorded by Riffo. Despite the wide geographical distribution of H. macrops, species richness in this host does not increased towards lower latitudes, as proposed by Rohde (1992, 1993) and Scott (1982). Replacement of species of parasites does occur and the structure of component communities differ markedly compared to those of Riffo (1991). In the central-southern area the infracommunities of H. macrops show predominance of ectoparasites. The most important species are the monogenean Neoheterobothrium sp. (erroneously identified as Hargicotyle sp., in Riffo 1991) with 96 % prevalence and the copepod, L. edwardsii with 68% prevalence. In our study, the predominant species were Neoheterobothrium sp. (71% prevalence) and the endoparasites, Floridosentis sp. and C. australe with prevalences of 52% and 55% respectively. The parasite fauna of the flatfish Paralichthys adspersus from northern Chile (23ºS) shows higher species richness, 22 parasite species (Oliva et al. 1996). Six of these are shared with H. macrops (E. hippoglossi, N. surmenicola, S. pleuronectis, C. australe, and Anisakis sp.). Even though P. adspersus has a greater species richness only two species, the digenean Cainocreadium sp. and the larval acanthocephalan, C. australe with prevalences of 54% and 56% respectively predominate (Oliva et al. 1996). The generic composition of the parasite fauna of H. macrops is similar to species of hosts (Pleuronectiformes) from other geographical localities sharing the genera Entobdella, Neo-heterobothrium, Corynosoma, and the complex eucestode S. pleuronectis. The species richness in H. macrops is lower when compared with other species of flatfish from the Northern Hemisphere (Table II), mainly due to the almost complete absence of digeneans and the scarcity of cestode and nematode species in H. macrops. The lower species richness in H. macrops has also been observed in other species of fish from the northern Chilean coast (Oliva et al. 1996, González & Acuña 1998, Oliva 1999). Sixty percent of the parasite fauna of H. macrops is composed of parasites with a low specificity (H. chilensis, Gliptonobdella sp., C. australe, S. pleuronectis, N. surmenicola, Anisakidae). Only the monogenean genera Neoheterobothrium and Entobdella are specific for flatfishes. The increase in intensity of Neoheterobothrium sp. with host age could be a consequence of a long exposure time to the infective stage of this monogenean and probably also to a larger gill surface in older, larger fishes. Likewise, the increase in prevalence of Neoheterobothrium sp. and H. chilensis with host age could be explained by the habit of this host. The occurrence of older demersal fish species inhabiting deeper waters than their younger conspecifics has been repeatedly observed in the marine and freshwater environments (Macpherson & Duarte 1991). H. macrops, like other flatfishes lives close associated with the bottom, the sedentary habit and relative high population density of mature specimens at deeper waters (Villarroel et al. 2000), can be synergistic forces that increase the infection, at least for those species with a direct life cycle. Host population density and schooling behavior are important factors in providing the opportunity for larval monogenean (oncomiracidia) to have access to new hosts (Campbell 1983). H. macrops shows a higher population density from 150 to 350 m depth (Acuña et al. 1999) where the specimens used in this study were obtained. Moreover, the changes in the ectoparasite fauna with host age suggest that this flatfish lives in different habitat depths during its lifetime, thus being subject to different parasite infections and rates. Changes in the endohelminth fauna during host life history can be related to its feeding habits (Campbell et al. 1980). The decrease of prevalence and intensity of Floridosentis sp. may be related to change in the diet of H. macrops (Villarroel & Acuña 1999). They reported that young fishes (< 3 years) fed mainly on juvenile squat lobsters (Cervimunida johni and Pleuroncodes monodon), while fishes > 3 years fed mainly on the deep sea shrimp (Heterocarpus reedi). It is also possible that the decrease in prevalence and intensity of endohelminth may be related to fish migration to deeper waters where the intermediate host can be absent. Amphipods have been considered as intermediate hosts for acanthocephalans but have not been found as a dietary component of the bigeye flounder (Villarroel & Acuña 1999). However, crustaceans other than amphipods also harbor larval acanthocephalans (Oliva et al. 1992, Pulgar et al. 1995). The predominance of acanthocephalans in this fish can be suggested as a consequence of its food preference, basically crustaceans like juvenile squat lobsters and deep-sea shrimp. Since information about the life cycle of Floridosentis sp. is not available, we can only suggest as a hypothesis that the crustaceans are the intermediate hosts for acanthocephalans parasitizing H. macrops. Acanthocephalans have been considered the most rarely found helminth parasites of fishes (Campbell 1983), but this does not apply to H. macrops or related flatfishes from other geographical areas, since all of them show the presence of adult acanthocephalans (mainly Echinorhynchus gadi) and species of the genus Corynosoma. In the bigeye flounder, as much as 65% of the abundance are larval and adult acanthocephalans. Parasites may infect differently both sexes, because male and female fish often have different feeding habits (Rhode 1993). In our study, host sex did not have a significant influence on the parasite fauna of H. macrops, suggesting that habitat use and diet are similar for both sexes of this species. This agrees with the findings of Villarroel (pers. commun.), who did not detect spatial bathymetric segregation between males and females. Noble (1973) and Campbell et al. (1980) have indicated that benthic fishes frequently harbor digeneans and nematodes, but this did not occur in the bigeye flounder. Only 1.6% and 7% of the fishes were parasitized with species representing those parasite groups, respectively. The scarcity of these parasites in the bigeye flounder could be explained by the smaller numbers of intermediate hosts such as benthic crustaceans and mollusks. The parasite success in the deep sea is directly related to the abundance and diversity of hosts in the benthic community (Campbell et al. 1980, Oliva & Luque 1998), therefore, the parasite richness is correlated with the diversity of free-living fauna in a certain area (Poulin 1995). The narrowness of the continental shelf in this geographical area has also been considered as an important abiotic factor that explains this phenomenon (Oliva 1999). Thus, a combination of biotic and abiotic factors unique to the coast of Chile may explain the paucity fauna of trophically transmitted parasites, specifically digeneans, en H. macrops. ACKNOWLEDGEMENTS To research assistants who collected specimens onboard and to Lianella Díaz for collaboration in collecting parasites. REFERENCES
Copyright 2001 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc01203t2.jpg] [oc01203t1.jpg] |
| |||||||||

{kind=link}
{kind=link}