 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
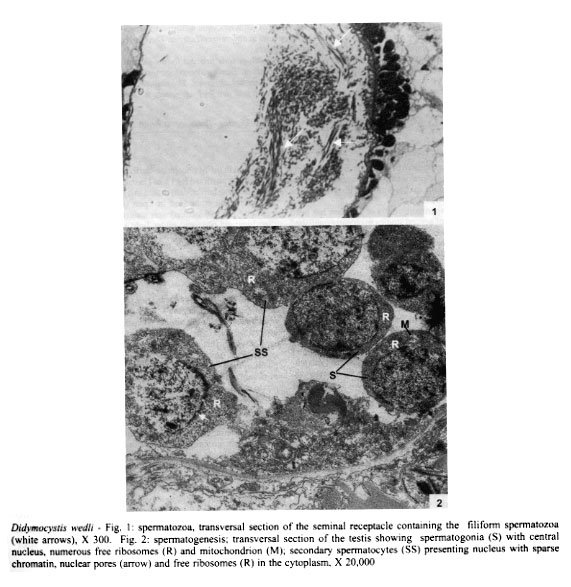
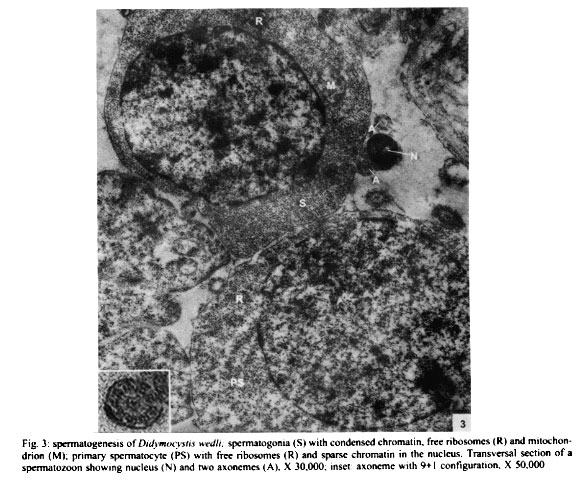
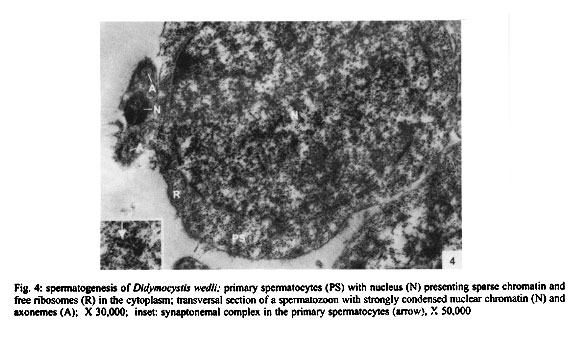
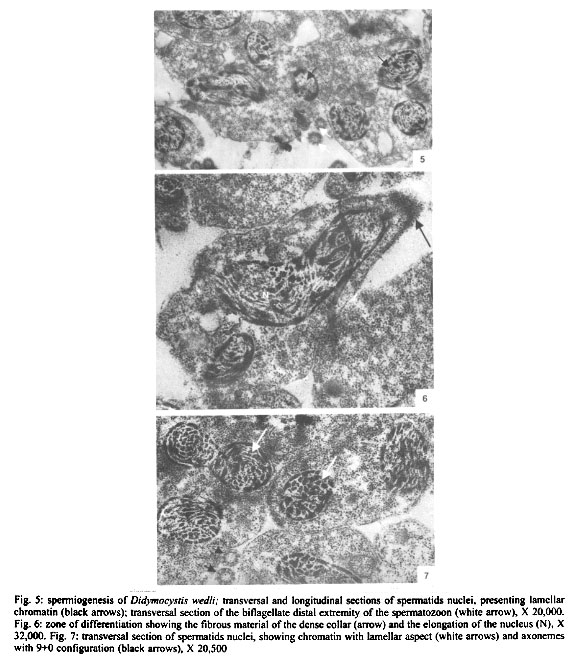
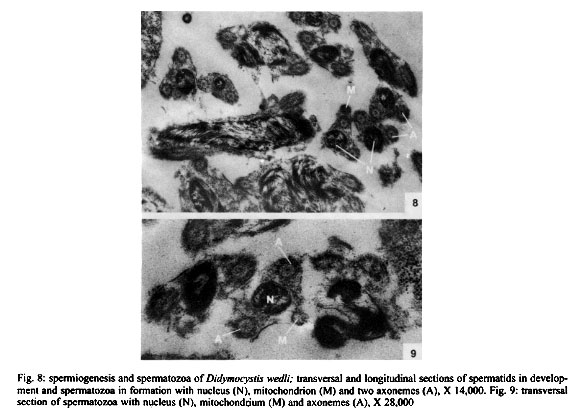
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 96(8) 2001, pp. 1153-1159 Spermatogenesis and Spermiogenesis in Didymocystis wedli Ariola, 1902 (Didymozoidae, Digenea) MC Pamplona-Basilio+, MFD Baptista-Farias, A Kohn++ Laboratório de Helmintos Parasitos de Peixes, Departamento de Helmintologia, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil Received 14 November 2000 Code Number: oc01220 The ultrastructure of the male reproductive system of Didymocystis wedli was studied for the first time, demonstrating spermiogenesis and spermatogenesis at different cell stages. The spermatozoa morphology was compared with that of other Digenea species. It was observed that the different cells of the spermatogenesis process follow the classic pattern reported for the majority of the parasitic platyhelminthes. During spermiogenesis, rootlet fibers, electrondense bodies and median cytoplasmic process were not observed. The mature spermatozoa of D. wedli were filiform, presenting nucleus, mitochondrion and two 9+1 axonemes, with a biflagellate distal extremity. Key words: spermatogenesis - spermiogenesis - Digenea - Didymocystis wedli The ultrastructure of spermiogenesis and spermatozoon have previously been used as characters for phylogenetic studies within the Platyhelminthes (Justine 1991, 1995). Indeed spermatogenesis of Digenea has been the subject of several studies in recently published papers, although these dealt with only a few of the existing species, such as Mesocoelium monas by Iomini et al. (1997), Echinostoma caproni by Iomini and Justine (1997), Postorchigenes gymnesicus by Gracenea et al. (1997), and Sac-cocoelioides godoyi by Baptista-Farias et al. (2000). In the family Didymozoidae, only two species have been studied: the ultrastructure of spermatozoon and spermiogenesis of Gonapodasmius sp. (Justine & Mattei 1982) and of Didymozoon sp. (Justine & Mattei 1983, 1984). The present study deals with the processes of spermatogenesis, spermiogenesis and the spermatozoon in a species of the family Didymozoidae, Didymocystis wedli Ariola, 1902, a parasite of Thunnus albacares recovered from the coast of the State of Rio de Janeiro, Brazil. MATERIALS AND METHODS D. wedli was collected from the gills of T. albacares from the coast of Rio de Janeiro. Worms were fixed in 0.1 M phosphate-buffered 2.5% glutaraldehyde, postfixed for 1 h in 1% osmium tetroxide in the same buffer, dehydrated in a graded ethanol series, and embedded in Epon (Luft 1961). Sections (0.5 µm thickness) were stained with toluidine blue and observed under light microscope. Ultrathin sections were collected on copper grids, double-stained with 2% alcoholic uranyl acetate and lead citrate (Reynolds 1963), and observed in a Zeiss EM 900 electron microscope. RESULTS Using TEM, it was observed that the testis rests on the basal lamina and are surrounded by a tunic of conjunctive tissue, forming a continuous layer around the cells in development (Fig. 2). Spermatogonia are cells which present a central nucleus, usually spherical, occupying almost all the cytoplasm. Nuclei contain irregular masses of strongly condensed heterochromatin. The cytoplasm contains several free ribosomes grouped in the format rosettes and mitochondria with slightly dense matrix (Figs 2, 3). Spermatocytes arise via mitotic divisions from spermatogonia; the nucleus presents granular nucleoplasm containing sparse chromatin and nuclear membrane with pores. The cytoplasm of these cells presents mitochondria dispersed throughout it and free ribosomes (Figs 2, 3, 4). The synaptonemal complex was observed at the primary spermatocytes; these structures are formed by the pairment and fusion of homologous chromosomes during meiosis (Fig. 4, inset). Mitochondria are similar to those observed in the spermatogonia. The spermatocytes divide and give rise to spermatids from which will originate the mature spermatozoon. The young spermatids are cells that present nuclei smaller than those seen in the other cells of the spermatogenesis process, presenting dense matrix with chromatin distributed in patches or in the form of dense filaments. This cellular stage is well characterized by the observation of mitochondria occupying a perinuclear position and free ribosomes. During the process, the spermatids differentiate, originating a projection at one of the edges, forming a collar of dense material through the appearence of a row of microtubules, characterizing a region called zone of differentiation (ZD) (Fig. 6). The nucleus elongate, becoming cylindrical, demonstrating the begining of the spermiogenesis process (Figs 5, 6). At this time, the mitochondria located at the perinuclear regions of young spermatids, migrate to the ZD. The chromatin modify, becoming strongly condensed and presenting a lamellar aspect (Figs 5, 6, 7). The nucleoli remain visible for a short period of time and are observed as dense bodies between the chromatin lamellae. Rootlet fibers, electrondense bodies and median cytoplasmic process were not visualized. At the end of the spermiogenesis process, the ZD narrows, the collar membrane approaches and each spermatozoon is released from the residual mass of cytoplasm. The mature spermatozoon is filiform (Fig. 1), with nucleus, mitochondrion and two axonemes located at the longitudinal axis, dividing into two flagellae at distal extremity (Figs 5, 8, 9). Peripheral microtubules were not observed. Each axoneme is composed of a central unit, surrounded by nine doublets of microtubules, characterizing the 9+1 configuration (Fig. 3, inset). In some cross sections, axonemes were observed with a 9+0 configuration, that can be explained by the absence of central pair in the posterior extremity of the spermatozoon, where the axonemes are separated (Fig. 5). DISCUSSION The process of spermatogenesis and spermiogenesis and the structure of the spermatozoon in D. wedli follow the same pattern described for other digenean species with some differences: during the spermiogenesis process, the fused spermatids in a common cytoplasmatic mass produce the ZD, and an elongation of the whole cell takes place, including nucleus, mitochondria and axonemes. The ZD and flagella fuse together, producing a mature spermatozoon. The ultrastructure of the spermatozoa of D. wedli presents peculiarities that clearly differentiate it from other digenean spermatozoa. According to Justine (1998) the incorporation of the axonemes into the cell occurs via proximo-distal fusion, but in D. wedli this fusion was not observed. During the spermiogenesis process of D. wedli, intercentriolar bodies, rootlet fibers, membrane arching and the median cytoplasmatic process are absent. Didymozoon sp. (by Justine & Mattei 1983) also does not present those structures, as well as the ZD, which was observed in the present study. In Gonapodasmius sp. (by Justine & Mattei 1982) only intercentriolar bodies and rootlet fibers are absent during the spermiogenesis. The majority of the Platyhelminthes display an homogeneous pattern for the spermatozoa with the nucleus, mitochondrion, two axonemes with a 9+1 configuration and cortical microtubules (Justine 1999). The spermatozoa of D. wedli follows this classic pattern, with the exception that the cortical microtubules are absent. Discrepancies occur within the group: the two axonemes of the spermatozoon in Didymozoon sp. (Justine & Mattei 1983) and the single axoneme of Schistosoma mansoni (by Kitajima et al. 1976) present a 9+0 configuration, representing the only exceptions within the Platyhelminthes with this configuration. Another species of Didymozoidae already studied, Gonapodasmius sp., shows the basic pattern for the structure of the spermatozoon. In the digeneans M. monas (by Iomini et al. 1997) and E. caproni (by Iomini & Justine 1997), one of the axonemes is longer than the other. According to these authors, the relative length of axonemes is a useful character for phylogeny of this group of parasites, as is the number of axonemes which have already been used for evolutionary studies of Monogenea (Justine 1991). The spermatozoa of parasitic platyhelminthes usually presents cortical microtubules, as in Gonapodasmius sp. (by Justine & Mattei 1982), while in Didymozoon sp. (Justine & Mattei 1983) and in D. wedli (present study) there are no microtubules. The data obtained in the present study may contribute towards elucidating the phylogeny of the family Didymozoidae, given that differences in the processes of spermiogenesis and in ultrastructure of the spermatozoon were observed in members of the same family. ACKNOWLEDGEMENTS To Antonia Lúcia dos Santos, Departamento de Helmintologia, Instituto Oswaldo Cruz (IOC-Fiocruz), for providing the fishes; to Dr Ortrud Monika Barth, Departamento de Virologia, IOC-Fiocruz, for the facilities offered and for the use of the electron microscope; and to Dr Simone Chinicz Cohen, Departamento de Helmintologia, IOC-Fiocruz, for her critical review of this manuscript. REFERENCES
+Corresponding author. Fax: +55-21-2260.4866.
E-mail: mclara@ioc.fiocruz.br Copyright 2001 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc01220f1-2.jpg] [oc01220f8-9.jpg] [oc01220f3.jpg] [oc01220f5-7.jpg] [oc01220f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}