
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 1, 2002, pp. 3-24
|
Mem Inst Oswaldo
Cruz, Rio de Janeiro, Vol. 97(1) 2002, pp. 3-24
A Critical Review
on Chagas Disease Chemotherapy
José Rodrigues Coura/+,
Solange L de Castro*
+Corresponding author. Fax:
+55-21-2280.3740. E-mail: coura@ioc.fiocruz.br
Departamento de Medicina Tropical
*Departamento de Biologia Celular e Ultraestrutura, Instituto Oswaldo Cruz-Fiocruz,
Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
This research was supported by grants
from the Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq), Programa de Apoio à Pesquisa Estratégica em Saúde
(Papes/Fiocruz), Fundação Carlos Chagas Filho de Amparo à
Pesquisa do Rio de Janeiro (Faperj), and Fundação Nacional da
Saúde (Funasa).
Received 5 November 2001
Accepted 10 December 2001
Code Number: oc02002
In this "Critical Review"
we made a historical introduction of drugs assayed against Chagas disease beginning
in 1912 with the works of Mayer and Rocha Lima up to the experimental use of
nitrofurazone. In the beginning of the 70s, nifurtimox and benznidazole were
introduced for clinical treatment, but results showed a great variability and
there is still a controversy about their use for chronic cases. After the introduction
of these nitroheterocycles only a few compounds were assayed in chagasic patients.
The great advances in vector control in the South Cone countries, and the demonstration
of parasite in chronic patients indicated the urgency to discuss the etiologic
treatment during this phase, reinforcing the need to find drugs with more efficacy
and less toxicity. We also review potential targets in the parasite and present
a survey about new classes of synthetic and natural compounds studied after
1992/1993, with which we intend to give to the reader a general view about experimental
studies in the area of the chemotherapy of Chagas disease, complementing the
previous papers of Brener (1979) and De Castro (1993).
Key words: Chagas disease - chemotherapy
- review - Trypanosoma cruzi - drug development - clinical treatment
BACKGROUND
Chagas disease is endemic in Latin
America, affecting 16-18 million people, with more than 100 million exposed
to the risk of infection (WHO 1997). Its etiological agent is Trypanosoma
cruzi, an hemoflagellate protozoan (family Trypanosomatidae, order Kinetoplastida)
(Hoare & Wallace 1966), whose life cycle involves obligatory passage through
vertebrate (mammals, including man) and invertebrate (hematophagous triatomine
bugs) hosts, in a series of stages. The trypomastigote ingested by the insect
differentiates into the proliferative epimastigote form that, on reaching the
posterior intestine, evolves to metacyclic trypomastigotes. This latter form,
following invasion of vertebrate host cells, undergoes differentiation into
amastigotes, which after several reproductive cycles transform to trypomastigotes,
the form responsible for the dissemination of the infection. The transmission
of the disease occurs mainly by the vector (80 a 90%), blood transfusion (5
a 20%) and congenital routes (0.5 a 8%) (Dias 2000).
In humans, after infection and a
subsequent incubation period, the acute phase of Chagas disease begins, and
in the absence of specific treatment, the symptoms persist for about two months,
with a mortality rate of 2 to 8%, especially among children. T. cruzi
is able to invade and multiply within different host cells, including macrophages,
smooth and striated muscles, fibroblasts and even neurons. The first reaction
to T. cruzi is focal mononuclear inflammation due to rupture of parasitized
cells. Within days to two weeks can be detected in the serum the presence of
immune complexes, decrease in C3 level, besides necrosis in the inflammatory
foci. Severe inflammation is usually accompanied by necrosis of parasitized
and non-parasitized cells, especially in the heart. Platelet aggregation, eosinophils
degranulation, microvascular pathology, edema, thrombosis, blood stasis and
ischaemia have also been demonstrated (Andrade & Andrade 1999).
After the acute infection, the patient
presents strong evidence of immunity, but has a tendency to remain infected.
Some parasites evading the immune response and focal inflammatory lesions are
seen in several organs. Amastigote forms can be detected by conventional histology,
by immunofluorescence and genomic markers by in situ hybridization. The
combination of rising immunity against the parasite with specific immunological
suppression of hypersensibility and reduction of inflammatory reaction seem
to be the main pathways to the indeterminate phase of Chagas disease (Andrade
1999).
In the chronic phase that follows,
most patients remain asymptomatic, with about 20 to 50% of the cases, accordingly
to the endemic area analyzed, developing the characteristic symptoms of this
phase, namely cardiac, digestive or neurological disturbances (reviwed in Brener
et al. 2000). Chronic, active, fibrosing myocarditis have been attributed to
hypersensibility to parasite antigens, neoantigens or autoimmunity. The presence
of cross-reactive antigens between heart muscle and T. cruzi has been
demonstrated, but the autoimmunity does not entirely explain Chagas heart disease.
The high positivity of xenodiagnosis and hemoculture, and reactivation of chronic
disease by immunosuppression demonstrate the presence of the parasite in chronic
cases. High frequency of parasites and/or antigens associated with myocardial
inflammation is an important guide to the therapeutic procedures in the chronic
phase.
The pathogenesis of Chagas disease
is not yet completely defined and understood, with two basic inflammatory lesions,
one focal and the other diffuse. The focal lesion is associated with the parasite,
and occurs when parasitized cells are disrupted. The diffusion lesion is out
of proportion in relation to the presence of parasites. During the acute infection
there are diffuse lesions in the heart and focal lesions in several other organs.
In the chronic heart disease, severe fibrosis and inflammatory lesions seem
not to be related only to the presence of T. cruzi, but also to a strong
delayed hypersensibility host response and ischaemic lesions (Andrade 1999,
Andrade & Andrade 1999, Higuchi 1999). Two mechanisms are proposed for pathogenesis
in chronic chagasic infections: the persistence of the parasite results in chronic
inflammatory reactivity and it induces immune responses targeted at self-tissues
(reviewed in Tarleton 2001). Several clinical reports reinforces the first mechanism
(Higuchi et al. 1993, 1997, Jones et al. 1993, Anez et al. 1999), while the
main support of the second one is that signs of the disease are evident in tissues
in the apparent absence of parasites.
Several reviews about clinical and/or
experimental Chagas disease treatment have been published as articles (Coura
& Silva 1961, Prata 1963, Brener 1975, 1979, 1984, De Castro 1993, Coura
1996, Urbina 1999, Stoppani 1999) and as book chapters (Brener 1968, 2000, Cançado
& Brener 1979, Cançado 1968, 1985, 1997, 1999, 2000, Rassi &
Luquetti 1992, Storino et al. 1994). We will detail here just some of the publications
and refer readers to the reviews cited above.
The aim of this "Critical Review"
is to analyse drugs employed in the clinics since the 70s bringing attention
to the recommendations about treatment, and evaluation of cure, and the studies
about the development of new drugs, considering potential targets in the parasite
and summarizing the experimental studies performed with new compounds assayed
against T. cruzi after 1992/1993. For a more complete coverage of experimental
in vitro and in vivo studies we suggest the reviews of Brener (1975, 1979) and
De Castro (1993).
EXPERIMENTAL
AND CLINICAL TREATMENT
Drugs assayed up to the decade
of 70
The first compounds assayed experimentally
for the treatment of Chagas disease, soon after its discovery by Carlos Chagas
in 1909, were atoxyl (arsenical), fucsin (rosanilin dye), tartar emetic (or
antimony potassium tartarate, a pentavalent antimonial) and mercury chloride,
employed experimentally by Mayer and Rocha Lima (1912, 1914) and all of them
without effective results. Until the publication of the "Manual de Doenças
Tropicaes e Infectuosas" by Carlos Chagas and Evandro Chagas (1935) "there
was no specific treatment for American trypanosomiasis. Drugs with trypanocidal
activity have been assayed by a great number of researchers, but without success",
affirm the authors (p. 144).
Among the chemotherapeutic agents
employed until 1962 stand out quinolein derivatives, several other antimalarials,
arsenobenzoles, and other arsenicals, phenantridines, salts of gold, bismuth,
copper and tin, sodium iodide, gentian violet, aminopterin, para-amino salicylic
acid, nicotinic acid hydrazide, antihistaminics, sulfonamides, ACTH, cortisone,
stylomycin derivatives, amphotericin B, more than 30 antibiotics and some nitrofurans
(reviewed in Coura & Silva 1961, Brener 1968, Cançado 1968).
Brener (1968) made a meticulous evaluation
of the experimental drugs assayed in vitro and in vivo against T. cruzi,
registering 27 compounds and more than 30 antibiotics, assayed between 1912
and 1962, that were inactive. He also considered that the following compounds
had a suppressive effect on the parasitemia but were not curative: the bisquinaldine
Bayer 7602 (Cruzon, Imperial Chemical Industry, UK), the phenatridine carbidium
sulfate (74C48), aminoquinolines (pentaquine, isopentaquine and primaquine),
trivalent arsenicals (Bayer 9736 and Bayer 10557 also named spirotrypan), aminoglycoside
of stylomycin, nitrofurans and antibiotics.
Packchanian (1952, 1957) opened a
new and promising line of potential drugs with the nitrofurans that led to nitrofurazone
(5-nitro-2-furaldehyde-semicarbazone). This derivative administered by oral
route for 53 days consecutively in the dose of 100 mg/kg/day mice experimentally
infected with T. cruzi cured 95.4% of the animals (62/65) (Brener 1961)
Ferreira (1961, 1962) and Ferreira
et al. (1963) treated the first ten cases of acute Chagas disease with this
nitrofuran, obtaining "good clinical results" with few collateral
effects but the xenodiagnosis became positive in five cases after treatment
Coura et al. (1961, 1962) treated 14 chronic cases with this drug in long-term
schemes, observing in the first four patients, that received progressive doses
of 10 to 30 mg/kg/day, important side effects that led to suspension of the
treatment due to severe sensitive polyneuropathy, that began at the third week
of nitrofurazone administration. With reduction of the dose to 10 mg/kg/day
and association with complex B, administered by parenteral route, five patients
tolerated the treatment for 60 days, in spite of the side effects (anorexia,
weight loss, paresthesia, and sensitive polyneuropathy). Another patient was
treated with 20 mg/kg/day and presented paresthesics manifestations only at
the 53th day of treatment, evolving to a severe sensitive polyneuropathy, with
termination of the treatment by his own decision (informed consent). From the
six patients submitted to long-term treatment, two of them were considered cured,
based on xenodiagnosis and serology (complement fixation) persistently negative.
Cançado et al. (1964) also treated five chronic patients with 10 mg nitrofurazone/kg/day
during 10 to 34 days when the treatment had to be suspended due to polyneuritis,
being all the five patients considered as therapeutic failures.
In a critical analysis of the literature
about the clinical experiences, Cançado (1968) emphasized the lack of
methods in the execution, preferential selection of acute cases, based on the
remission of the symptoms and signs that could also be spontaneous. He cited,
as example, the reports of Mazza et al. (1937, 1942) and Pifano (1941) about
the results with Bayer 7602 and Bayer 9736 that were considered "excellent
results" only based on the reduction of the symptoms and signals. Both
compounds were in fact ineffective, since the xenodiagnosis after treatment
remained positive and untreated cases had also reduction of the symptomatology.
In subsequent works (Cançado et al. 1973, Cançado 1981) reviewed
the results of therapeutic trials in the period of 1936 to 1965 and proposed
basic criteria for the evaluation of the specific treatment, that were later
updated by 15 experts from Latin America (OPS/OMS 1998).
Following the requirements of the
World Health Organization (WHO) the ideal drug for the treatment of Chagas disease
should fulfill the following requirements: (i) parasitological cure of acute
and chronic cases; (ii) effective in single or few doses; (iii) accessible to
the patients, in other words, of low cost; (iv) no collateral or teratogenic
effects; (v) no need of hospitalization for the treatment; (vi) no induction
of resistance.
As we will see bellow this ideal
drug does not exist and possibly it will take a long period of time to be obtained.
Since the end of 1960 beginning of the 70s two drugs have been used for the
treatment of Chagas disease: nifurtimox and benznidazole.
Nifurtimox and benznidazole
The drugs and Chagas disease treatment
Nifurtimox (Nif) is a 5-nitrofuran
(3-methyl-4-(5'-nitrofurfurylideneamine) tetrahydro-4H-1,4-tiazine-1,1-dioxide
(Bayer 2502) and benznidazole (Bz) is a 2-nitroimidazole (N-benzyl-2-nitroimidazole
acetamide (RO 7-1051), commercialized, respectively, with the names Lampit and
Rochagan in Brazil (Radanil in Argentina) (Fig.
1a,b). Nif that
was the most active 5-nitrofurfurilidene derivative experimentally assayed (Bock
et al. 1969) and Bz showed a high in vitro and in vivo activity against T.
cruzi (Richle 1973). Since the 80s Nif had its commercialization discontinued,
first in Brazil, and then in Argentina, Chile and Uruguay. The mode of action
of Nif involves generation of nitroanion radical by nitroreductases that, in
the presence of oxygen, led to reactive intermediates and being T. cruzi
is partially deficient in free radical detoxification mechanisms, it is susceptible
to such intermediates (reviewed in DoCampo & Moreno 1986). On the other
hand, this oxidative damage was not the key action of Bz, the detection of corresponding
nitroanion radical occurred only at concentrations much higher than those that
killed the parasite. The action of Bz could involve covalent bond or other interactions
of nitroreduction intermediates with parasite components (Polak & Richle
1978), or binding to DNA, lipids and proteins (Diaz de Toranzo et al. 1988).
Nif and Bz have been employed by
different authors, especially in Brazil, Chile and Argentina (Cançado
et al. 1969, 1973, 1975, Cançado & Brener 1979, Bocca-Tourres 1969,
Rubio & Donoso 1969, Schenone et al. 1969, 1972, 1975, 1981, Rassi &
Ferreira, 1971, Rassi & Luquetti 1992, Cerisola et al. 1972, 1977, Prata
et al. 1975, Ferreira 1976, 1990, Coura et al. 1978, 1997, Macêdo &
Silveira 1987, Viotti et al. 1994, Andrade et al. 1996, Sosa Estani et al. 1998).
The results obtained with both drugs
varied according to the phase of Chagas disease, the period of treatment and
the dose, the age and geographical origin of the patients. Good results have
been achieved in the acute phase, in recent chronic infection (children under
12 years old), congenital infection and laboratory accidents. For the acute
phase treatment and congenital cases it is recommended 8 to 10 mg/kg/day of
Nif or 5 to 7.5 mg/kg/day of Bz during 30 to 60 days consecutively, and divided
in two or three daily doses. Patients with less than 40 kg can take up to 12
mg/kg/day of Nif and up to 7.5 mg/kg/day for Bz during 30 to 60 days (OPAS/OMS
1998). For recent chronic infection (children under 12 years old) or individuals
infected in the last 10 years, the treatment should be made with 8 mg/kg/day
of Nif or 5 mg/kg/day of Bz during 30 to 60 days. In the case of accidental
infection the treatment must begin immediately and last for only 10 to 15 consecutive
days. Cases of late chronic infections without clinical manifestation or with
mild cardiac or digestive manifestations should be treated during 60 to 90 days,
in accordance with the tolerance to the drugs, aiming to prevent or reduce the
evolution of Chagas disease to more severe forms, a fact that is not yet definitely
proved.
Side effects and contraindications
The more frequent collateral effects
with Nif treatment are anorexia, loss of weight, psychic alterations, excitability
or sleepiness and digestive manifestations, such as nausea, vomit and occasionally
intestinal colic and diarrhea. The adverse reactions with Bz could be classified
in three groups: (i) symptoms of hypersensibility, dermatitis with cutaneous
eruptions (usually appearing between the 7th and 10th day of treatment), generalized
edema fever, lymphadenopathy, articular and muscular pain; (ii) depression of
bone marrow, thrombocytopenic purpura and agranulocytosis, the most severe manifestation;
(iii) polyneuropathy, paresthesia and polyneuritis of peripheric nerves.
The two most serious complications
induced by Bz are agranulocytosis, initiated by neutropenia, sore throat, fever
and septicemia, and thrombocytopenic purpura, characterized by reduction of
platelets, petechiae, hemorrhagic blister and even mucosal bleeding. At the
first signs of such manifestations, medication must be immediately suspended
and should be started a treatment with antibiotics in the case of septicemia
plus corticosteroids for the control of the agranulocytosis and of the thrombocytopenic
purpura. Other manifestations of intolerance and hypersensibility could be circumvented
with the reduction of the dose or suspension of the drug, depending on their
intensity, introduction of symptomatic medication and eventually of anti-histaminics
and corticosteroides.
Teixeira et al. (1990, 1994) have
been alerting for the appearance of lymphomas and mutagenic and carcinogenic
activities in experimental animals (rabbits and mice) treated with Bz. However,
a broad review of thousands of patients treated with these drugs by several
authors has not demonstrated such effects.
Bz and Nif should not be indicated
for pregnant patients, in cases of severe diseases associated with Chagas disease,
such as systemic infections, cardiac, respiratory, renal or hepatic insufficiency,
hemopathies and neoplasies without the possibility of treatment, old-aged and
very debilitated persons.
Effect of the treatment on the evolution
of Chagas disease
Since 1969 several therapeutic experiences
have been performed in acute and chronic cases of Chagas disease using Nif (Bocca-Tourres
1969, Rubio & Donoso 1969, Schenone et al. 1969, 1972, Rassi & Ferreira
1971, Cerisola et al. 1972, 1977, Silva et al. 1974, Prata et al. 1975, Cançado
et al. 1975, 1976, Cançado & Brener 1979), Bz (Schenone et al. 1975,
Ferreira 1976, Coura et al. 1978, Viotti et al. 1994, Andrade et al. 1996, Sosa
Estani et al. 1998) and also comparing the efficacy and tolerance of both drugs
in different groups of patients, therapeutic schemes and periods of follow-up
and cure evaluation criteria (Schenone et al. 1981, Ferreira 1990, Coura et
al. 1997, Lazzari & Freilig 1998).
The results of such experiences showed
a great variability, according to the authors, and the type of casuistic and
of cure control employed. In general, results obtained were good for acute phase
and recent infection cases, especially among children, who, besides tolerating
a long-term treatment much better, presented high indexes of cure, as demonstrated
in a randomized field study with Bz in Brazil (Andrade et al. 1996) and in Argentina
(Sosa Estani et al. 1998). For acute cases and recent infections an average
index of parasitological cure around 60% is estimated. In relation to chronic
infection cases, results have been poor, for the Brazilian experience, while
in the South Cone, they were much better, possibly due to the type of parasite
strain (Silva et al. 1974, Cerisola et al. 1977).
Studies on the clinical evolution
of Chagas disease after specific treatment are controversial and results are
not convincing, due to differences in casuistics, methods of evaluation, time
of follow-up and interpretation of the data. Macêdo and Silveira (1987)
studying 171 adults with chronic disease treated with Nif or Bz with a follow-up
of 7 years, observed electrocardiographic (ECG) evolution of cardiopathy in
6.7% of the cases, against 8.8% for untreated patients, indicating no significant
differences between the two groups. Ianni et al. (1993) monitored 33 adults
in the indeterminate phase for 8 years, reported the evolution in 13.3% of the
cases treated with Bz (n = 15) and 0% of the cases that received placebo (n
= 18), not allowing a definitive conclusion, due to the small casuistic. Miranda
et al. (1994 apud OPAS/OMS 1998), in 120 patients (adults and children),
observed ECG evolution in 10.5% of those treated with Bz and 63.6% of the placebo
group. Although these authors monitored the patients for 10 to 16 years, the
combination of the data obtained with adults and children and the ECG interpretation
made the analysis of the results obtained difficult.
Viotti et al. (1994) in a well-designed
study with 201 adult patients monitored for 8 years, observed ECG evolution
in 7/131 cases (5.3%) treated with Bz (5 mg/kg/day for 30 days) and 16/70 (22.8%)
in the control group. In patients with more than 50 years old, the ECG alterations
occurred in 3/36 (8.3%) of the treated cases and 7/40 (17.5%) of the untreated
ones; differences were not statistically significant. For those under 50 years
old, such alteration occurred in 4/95 (4.2%) and 9/30 (30%), respectively, for
treated and untreated patients, and were significantly different. Two patients
died during the follow-up, one treated and one untreated. Although the study
has been well conducted, after 8 years of follow-up, 68% of the untreated patients
presented positive serology against 48.2% of the Bz-treated group. The low number
of ECG alterations and their frequent mutability in chronic cases make the interpretation
of the data difficult.
On the other hand, Fragata Filho
et al. (1995) reported, in a study with 120 chronic patients with follow-up
for 7-8 years, ECG evolution in 7% for Bz-treated cases (n = 71) and 14.3% for
the untreated group (n = 49).
Andrade et al. (1996) performed a
randomized field study in the State of Goiás (Brazil) treating children
between 7 and 12 years old with positive serology for Chagas disease. Sixty-four
patients received 7.5 mg/kg/day of Bz for 60 days and 65 received placebo. From
these 129 children, 88.7% (58 treated and 54 control) were monitored for three
years. The authors considered the treatment effective in 55.8% of the treated
cases (most of them with a significant decrease in serological titers). However,
no significant results were observed when the ECG abnormalities were compared.
In a similar way, in Salta (Argentina), Sosa Estani et al. (1998) treated 55
children from 6 to 12 years old with 5 mg Bz/mg/day for 60 days and 51 children
with placebo, and monitored the study for four years. They observed negativation
or significant decrease of the specific serology in 62% of the treated group
and none for the control group. In relation to xenodiagnosis the positivity
was 4.7% and 51.2%, respectively for treated and untreated children, indicating
an important suppressive activity of the treatment, however, the ECG alterations
were similar for both groups: 2.5% (1/40) and 2.4% (1/41).
Rules and recommendations for
the clinical treatment
A meeting of 13 specialists promoted
in Brasília by the Ministry of Health of Brazil summarized by Luquetti
(1997) and another one by the Pan American Health Organization and World Health
Organization (OPAS/OMS 1998) that took place in Instituto Oswaldo Cruz (Fiocruz)
in April 23-25 of 1998, established some rules and recommendations for the etiologic
treatment of Chagas disease, with the present available drugs, that are summarized
bellow:
Acute phase
In this phase the parasite is detected
by direct examination of peripheral blood by weat smear or in stained cover
slides or by concentration methods (centrifugation of the blood and microscopic
examination of leukocyte cream or stained quantitative buffy coat techniques).
With or without clinical manifestations, the detection of the parasite by direct
methods or determination of IgM levels allows the diagnostic of the acute phase.
Independently of the mechanism of transmission (vectorial, transfusional, by
oral route, or laboratory accident) the patients must be treated, as indicated
previously, since about 60% of them could be cured in the acute phase.
Congenital infection
The diagnostic of congenital infection
is based in cases of children from infected mothers, serologically positive,
who presented T. cruzi in the blood of umbilical cord, specific IgM in
the serum soon after birth, or IgG after 6 months, when the possibility of vectorial,
transfusional and oral transmission are absent. The treatment is similar to
that of the acute phase patients.
Accidental infection
The person, technician or researcher,
that working with T. cruzi, was accidentally punctured by infected needle,
ingested or had any contact with infected materials in lesions, wounds, mucosal,
or any other form indicative of the possibility of the parasite penetration
is considered infected. In these cases, blood is collected for serological test
and treatment immediately begins, during 10 to 15 days, repeating the serology
after 15, 30, and 60 days after the accident. It is recommended that all the
laboratories that work with T. cruzi have the drug available.
Chronic phase
Recent chronic phase - Recent
chronic phase is considered when the infection was acquired in the last 10 years,
including children up to 12 years old or adults that have been infected occasionally
in endemic areas of by blood transfusion in a known interval of time. The published
work indicates that the results obtained by the etiologic treatment in this
phase are much better than that performed in late chronic patients.
Late chronic phase - Patients
with more than 10 years of infection are considered late chronic cases. There
is no agreement about the clinical evolution of such cases, and parasitological
cure is obtained in 10 to 20% of the patients, according to different experiences.
The treatment must be elective, with priority to cases in the indeterminate
form or with minor pathology that may be monitored by long periods of time,
after treatment.
Transplant of organs - In
cases of transplants it is always necessary to perform specific serological
reaction both in the donor and the receptor. The transplant of organs from patients
with infection by T. cruzi could transmit the parasite to the receptor,
especially during the immunosuppression phase. On the other hand, in the infected
receptor a reactivation of the disease could occur through the immunosuppression,
compromising even the transplanted organ, mainly in cardiac cases. Jatene et
al. (1997) refer to reactivation of Chagas disease in 40% of cases derived from
immunosuppression in heart transplants. Both the donor and the receptor must
be treated with Bz in the dose of 5 mg/kg/day for at least 60 days. Tominori-Yamashita
et al. (1997) considered that "allopurinol seems a safe and effective treatment
for reactivated Chagas disease after heart transplantation, although it is not
recommended as a post-transplantation prophylaxis because reactivation of the
disease is unpredictable".
Reactivation of Chagas infection
Reactivation of the chronic disease
can occur due to immunosuppression by several diseases, such as leukemia, lymphoma
and other neoplasies, infection by HIV/AIDS and in the cases of transplants
with immunos-supression. Meningoencephalitis and acute myocarditis are the most
frequent manifestations (reviewed in Ferreira et al. 1997a). In chronic patients
parasites were detected in cutaneous lesions after transplant with immunosuppressive
treatment and in the smooth muscle after cancer chemotherapy. Ferreira et al.
(1997a,b) made an extensive review about this topic, recommending as first choice
the treatment with Nif or Bz, indicating as alternatives triazole derivatives
and allopurinol. Long-term secondary prophylaxis should be recommended for patients
who respond to therapy, although it is uncertain which drug to use for this
purpose (Ferreira et al. 1997b). Anyway all HIV-positive cases, patients with
neoplasies or candidates to transplants must be thoroughly investigated for
the possibility of concomitance with chagasic infection. There is no consensus
about the prophylatic indication of the etiologic treatment of the infection,
in cases without clinical reactivation, but with xenodiagnosis or hemoculture
positive for T. cruzi.
Patients in the chronic phase and
receiving corticoid because of concomitant diseases were treated with Bz since
the beginning of the use of corticoid (n=12) or 15 days afterwards (n = 6) (Rassi
et al. 1999). The authors observed that Bz prevented an increase of parasitemia,
and suggested that in immunocompromized patients with chronic Chagas disease
this drug could be useful.
Where and who should treat the patient
The patients in severe acute phase
and the congenital symptomatic cases with diagnostic at birth, must be hospitalized
for treatment. The acute oligosymptomatic or chronic cases can be treated in
basic health care units under the supervision of an experienced physician. Bz
and Nif must be considered drugs of high complexity, and recommended only by
professionals with solid information about the side effects and the disease
in itself. The acute phase and the accidental infection are emergency situations,
and treatment must start immediately even with a professional without experience
that must search a colleague or a qualified institution for orientation.
Evaluation of cure
The evaluation of cure of Chagas
disease is certainly the more complex aspect of its treatment, leading several
times to diverse and controversial results, in relation to both parasitological
cure and clinic cure. The term parasitological cure itself is of difficult interpretation
and the evaluation is almost impossible, since it would mean the total elimination
of the parasite not only from the blood but also from all tissues. So, in humans
it is not viable to be confirmed. On the other hand, clinic cure is the long-term
evaluation and several times uncertain due to the pathogenesis of the disease,
which involves the action of parasite and the immune and autoimmune response
of the patient and in antigenic complexes deposition, generation of antibodies,
inflammatory reactions, tissular lesions with cellular degeneration, ischaemia,
fibrosis and their consequent clinical manifestations, sometimes for long periods
of time.
For the evaluation of experimental
animals, mostly in drug assays, the situation is less complex. The in vitro
tests (tissue culture) are not necessarily reproduced in vivo. On the other
hand, as occurs in humans, a suppressive effect on the parasitaemia does not
correspond exactly to the effect of a drug in tissues. In his pioneer work Brener
(1961) demonstrated parasitological cure in 94.5% of the mice treated with nitrofurazone.
Later, Brener et al. (1969) demonstrated by electron microscopy that 13.5% of
the amastigotes of the Y strain were intact in heart cells of the treated animals.
A question remained: were these findings due to populations of the parasite
with primary resistance or the drug did not reach all the infected cells? The
group of Andrade reported development of resistance to both nitroheterocycles
and the influence of T. cruzi strain in the cure rate, for example, Bz
cured 87% of the mice infected with the Peru strain, and only 16.7% in the case
of Colombian strain (Andrade et al. 1975, 1977, Andrade & Figueira 1977).
Resistance to both drugs, including cross resistance, was also observed in animals
infected with the Y strain (Costa Silva et al. 1990). Could this be due to a
mechanism similar to that questioned above?
Parasitological evaluation
The suppressive activity on the parasitaemia
is almost immediate after the beginning of the treatment when the strain (population)
of T. cruzi is susceptible to the drug employed. In acute cases this
fact could be verified by the direct examination of blood in fresh or stained
preparations or by concentration methods. In chronic cases, the usual methods
are xenodiagnosis standardized with 40 nymphs of 3rd/4th stage of Triatoma
infestans or by 20 nymphs of this species and another 20 from Panstrongylus
megistus or of other species that could give similar yield. The nymphs are
distributed in four boxes (10 per box); two are placed in the internal face
of each arm for about 30 min (Coura et al. 1991). In this study we observed
a positivity of 50.7% in 570 xenodiagnosis performed in 246 patients. Nowadays,
due to ethical questions and comfort for the patients, the preferential method
is the artificial xenodiagnosis that consists in collection of 10 ml of the
blood that are placed in a condom type membrane, with external heating, and
then the nymphs are added and the reading done after 30, 45 and 60 days. The
yield obtained is similar to that of the natural test (Pineda et al. 1998).
In a multicenter study that involved
researchers from 10 Brazilian institutions and 312 Bz-treated patients monitored
by a media of 12 xenodiagnosis performed after treatment, suppression of parasitaemia
was demonstrated in 78% of the cases (Coura et al. 1978). In another comparative
controlled study with 77 chronic patients treated with Bz (n = 26), Nif (n =
27) or placebo (n = 24), suppression of the parasitaemia monitored xenodiagnosis
by one year after treatment was achieved in 98.1% (2/110) of the Bz group, and
in 90.4% (75/83) of the Nif group (Coura et al. 1997). However, this result
does not imply that cases with xenodiagnosis negative are cured since only 34.3%
of the control group was positive.
The hemoculture is the second parasitological
method of choice for the control of cure in chronic Chagas disease, being equivalent
in sensibility to xenodiagnosis (Chiari & Brener 1966). Both methods have
a tendency to increase the positivity with the number of tests performed, amount
of blood employed, cultivation medium, interval of time between blood collection
and cultivation and other factors emphasized by Chiari et al. (1989) aiming
standardization of the assay. Using 30 ml of blood seeded in six test tubes
with LIT medium and readings up to 60 days, Chiari et al. (1989) obtained a
positivity up to 50%, while with small modifications of the technique such as
direct seeding soon after blood collection, refrigerated centrifugation for
a short time, gentle homogenization and up to 120 days led Luz et al. (1994)
to a positivity of 94%, not achieved by any other author, for assays with chronic
cases of Chagas disease.
Polymerase chain reaction (PCR) was
a major advance for the parasitological control of the cure of Chagas disease,
with positivity 2 to 3 times higher for chronic cases when compared to routine
xenodiagnosis and hemoculture. By this technique it is possible to detect one
parasite or a fragment of T. cruzi DNA in 20 ml of blood (Ávila
et al. 1991).
Sturm et al. (1989) amplifying minicircles
of DNA of T. cruzi obtained fragments of 83 and 22 pairs of base (bp)
from variable regions that were employed for detection of the parasite (Moser
et al. 1989). Ávila et al. (1991) using a solution of 6 M guanidine plus
EDTA and equal amount of blood of chronic patients, promoted lysis of proteins
and maintained the integrity of the DNA sample at room temperature. The treatment
of this lysate with phenantroline-copper (OP-Cu2+) led to the cleavage
of DNA and liberation of minicircles, allowing the identification of a single
parasite in 20 ml of blood when three initiators for the fragments of 83, 122
and 330 bp from the variable and constant regions of the minicircles. By this
technique, the authors identified T. cruzi in samples of 10 ml of blood
of five chronic patients, four of them with negative xenodiagnosis.
Several authors (Wincker et al. 1994,
Britto et al. 1995, 2001, Junqueira et al. 1996) have demonstrated the efficiency
of PCR for the diagnosis of chronic disease and for the control of cure after
treatment. However, Junqueira et al. (1996), in a comparative study among PCR,
xenodiagnosis and hemoculture in 101 chronic cases, observed positivity of,
respectively, 59.4%, 35.6% and 25.7%, but in five cases with positive xenodiagnosis
and/or hemoculture, the result of PCR was negative. Recently, Britto et al.
(2001) comparing PCR and xenodiagnosis in the control of cure of 85 chagasic
patients submitted to specific treatment and 15 chronic assymptomatic cases
that received placebo, reported that in all the cases of positive xenodiagnosis,
positivity was obtained also by PCR. On the other hand when xenodiagnosis was
negative, PCR was positive in 18.5% of the acute phase group (n = 37), 29% of
the chronic phase group (n = 45) and 57.1% of the control group. These results
demonstrate the advantage of PCR over conventional techniques to demonstrate
persistent infections in patients that underwent chemotherapy.
Serological evaluation
This evaluation is certainly the
most simple, more broad and reliable for the control of cure of chagasic infection
after treatment, especially in the chronic phase when the serology is positive
in almost 100% of the untreated cases. Whereas the positivity of parasitological
methods depends on the random presence of the parasite in the blood sample,
the presence of antibodies is almost warranted in all the samples. On the other
hand, the serology since the Guerreiro and Machado reaction (1913) until the
qualitative and quantitative reaction of complement fixation (Kelser 1936, Freitas
& Almeida 1949), the indirect immunofluorescence assay (Fife & Muschel
1959, Camargo 1966), the hemagglutination assay (Neal & Miles 1970), the
conventional ELISA (Voller et al. 1975) and with recombinant antigens (reviewed
in Silveira 1992, Silveira et al. 2001), the lysis mediated by complement (Krettli
& Brener 1982, Kretlli et al. 1984) and finally techniques using immunoblots
have been improved as confirmatory tests due to their sensibility and specificity
(Umezawa et a1. 1996, 2001).
The three basic serological reactions
for the diagnosis of Chagas disease are indirect immunofluorescence, hemagglutination
and ELISA. During the years, the great polemics is their negativation in cases
of parasitological cure. Some authors as Cançado (1963, 1997) consider
cure as "definitive post-therapeutic reversion to negativity of parasitological
and serological tests", whereas others, like Rassi and Luquetti (1992),
Andrade et al. (1996) and Sosa Estani et al. (1998) admit a long period of negativation
of the reactions and even low serological titers as criteria of cure. Andrade
et al. (1991) demonstrated that, in mice infected with T. cruzi and parasitologically
cured by chemotherapy, parasite antigens persist in interstitial dendritic cells
in the spleen and the animals present positive serology. Recently Andrade et
al. (2000) reported the importance of the presence of parasite antigens in the
same type of cells in the heart of infected dogs and the pathogenesis of the
chagasic myocarditis probably by presentation of T. cruzi antigens to
immune-competent cells, and, as consequence, maintenance of the response to
the infection.
Clinical evaluation
This type of the cure evaluation
after chemotherapy is perhaps the most difficult and long topic to be addressed.
In this review, we have already discussed some aspects of the clinical evaluation
when analyzing the evolution of Chagas disease after treatment with Nif or Bz,
so, in this topic we will discuss only some essential clinical tests for monitoring
the disease, before, during and after treatment.
In the clinical evaluation of the
treatment we must consider, besides the anamnesis and clinical examination,
the electrocardiographic and radiologic aspects together with other non-invasive
tests with high sensibility, such as dynamic electrocardiography (Holter) for
the study of arrhythmias and echocardiography for the anatomophysi-ological
evaluation of the cardiac function, the endoscopy and manometry for the anatomofunctional
study of the digestive system and some other tests for evaluation of the autonomous
nervous system and neuronal lesion, besides biopsies for histological and histochemical
studies, that will not be evaluated here in depth.
A careful clinical examination after
a detailed anamnesis, especially analyzing the cardiovascular, digestive and
neurologic systems, before, during and after treatment, monthly in the first
year and at least once a year subsequently is fundamental importance for the
evolutive study of the patients. The ideal condition, when ethics allows, would
be the monitoring of a control group, of the same age and sex, untreated or
receiving a placebo, with aspect similar to the drug, for evaluation of collateral
effects and a comparative study between the treated and the control groups.
The standard electrocardiogram, with
the limb leads (D1, D2, D3, aVR, aVL and aVF) and chest leads from V1 to V6
is the most simple and most important examination in the clinical evaluation.
This test must be associated with the anamnesis and the physical examination
in each consultation during all the follow-up period.
The radiological examination is a
less sensible method and more expensive than ECG, so it should be performed
once before treatment, 6 and 12 months later and then once per year of monitoring.
This examination should consist of a chest RX with postero-anterior and lateral
views with esophagus contrasted immediately after ingestion of barium and also
after 1 min for the evaluation of the time necessary to drain the contrast (Rezende
et al. 1960). The barium enema with previous preparation and radiography of
colon is the only test capable of evaluating an established megacolon (Rezende
1959).
The dynamic electrocardiography and
the echocardi-ography are suitable techniques for evaluation of arrhythmias
and anatomophysiology of the heart and must be performed by a cardiologist.
In the same way, endoscopic and manometric tests need an experienced gastroenterologist.
The examination of the autonomous and peripheric nervous systems, with or without
stimulation with cholinergic drugs, such as pilocarpine, can be done by a physician
(Macêdo 1997).
The histopathology study of biopsies
fragments of the digestive system or endomyocardic with conventional microscopy
or analysis by immunoperoxidase or immunofluorescence are indicated only in
some research cases and could only be performed following strict protocols from
the ethics point of view.
New drugs in clinical tests
For several years in Brazil, and
more recently in Argentina, Chile and Uruguay, only Bz is commercialy available
as the development of drugs for tropical diseases is of little interest for
the pharmaceutical industry (Fairlamb 1999). After the introduction of Nif and
Bz, few compounds were assayed in chagasic patients.
Allopurinol
The results obtained with allopurinol
(4-hydro-xypyrazolo (3,4-d) pyrimidine HPP, Fig.
1c) in experimental animals and the knowledge about its mode of action led
to its clinical assays for the treatment of Chagas disease. This compound is
a hypoxanthine analog that acts as an alternative substrate of hypoxanthine-guanine
phospho-ribosyltransferase (HGPRT) and is incorporated into the RNA. This incorporation
leads to formation of non-physiological nucleotides and to blockade of the synthesis
de novo of purine nucleotides (reviewed in Marr 1991).
In the treatment of six acute phase
patients, allopurinol was ineffective, with maintenance of positive xenodiagnosis
and serology (Lauria-Pires et al. 1988). In a study, with chronic patients,
Galleano et al. (1990) treated two groups of patients with 600 and 900 mg/kg/day
of allopurinol for 60 days, compared with other two treated with Nif and Bz.
In the four groups the percentage of negativation of xenodiagnosis was in the
range of 75-92%, and those treated with allopurinol presented less collateral
effects. Allopurinol (600 mg/day for 2 months) was administered to two cases
of reactivation of Chagas disease due to cardiac transplant. Erythematous lesions
on the superior and/or inferior members characterized this reactivation. In
one patient the lesions disappeared in 3 weeks, and in the second one, after
2 weeks there was a clinical improvement of the lesions. In both cases after
treatment, xenodiagnosis and hemoculture tests were negative and in the follow-up
of 38 and 17 months, respectively, no reactivation of Chagas disease occurred,
even with continued immunosuppression (Tomimori-Yamashita et al. 1997). Apt
et al. (1998) treated 104 chronic patients with allopurinol (8.5 mg/kg/day for
60 days) that were monitored by clinical examination, serology, xenodiagnosis,
hemoculture and electrocadiogram. Parasitological cure was achieved in 44% of
the allopurinol-treated patients. The criteria for parasitological cure were
maintenance of negative xenodiagnosis and/or complement-mediated lysis for at
least four years. A double blind randomized longitudinal study must be performed
to reevaluate the efficacy of this drug for the treatment of Chagas disease.
Ketoconazole
In eight chronic patients, ketoconazole
(cis-(dl)-1-acetyl-4-[4-[[(2-2,4-dichlorophenyl)-2-(1H-imidazol-1-yl
-methyl)-1,3-ioxalan-4-yl]methoxy]-phenyl]-piperazine, Fig.
1d) was administered in doses between 3.1 and 8.7 mg/kg by oral route during
51 to 96 days and cure evaluation was performed by hemoculture, conventional
serology and complement-mediated lysis. The patients were monitored up to 60
months and it was observed that the drug was unable to erradicate the parasites,
from 6 out of 8 patients with positive hemoculture and two others with positive
serology (Brener et al. 1993).
In a case of reactivation of Chagas
disease in a patient in the inderteminate phase due to infection by HIV, ketonazole
(400 mg/day) was administered for 70 days leading to a negative xenodiagnosis.
The treatment was suspended by the patient´s own decision and after one
month occurred signs of reactivation of the disease, including development of
myocarditis. Bz was introduced (200 mg/day) and after four days, although negativation
of parasitemia, the patient presented signs of neurological deterioration. This
drug was maintained for 45 days and then replaced by ketoconazole (400 mg/day)
as a suppressive treatment. However, after a general clinical improvement, there
was a new neurologic deterioration and the patient died (Galhardo et al. 1999).
Ketoconazole was one of the first
imidazoles that showed in vitro activity against T. cruzi, with accumulation
of metabolites of sterol metabolism in epimastigotes. In vivo ketoconazole led
to parasitological cure in experimental animal in the acute phase, but was ineffective
in the chronic phase (reviewed in De Castro 1993). A synergic effect of ketoconazole
and Bz was observed in mice infected with the CL or Y strain, what did not occur
in the case of the Bz-resistant Colombian strain (Araújo et al. 2000).
Fluconazole and itraconazole
An haemophylic patient was infected
by blood transfusion with HIV and T. cruzi and brain biopsy revealed
the presence of amastigotes inside glial, macrophages and endothelial cells.
Initially he was treated with Bz (400 mg/day) but due to general worsening of
his clinical condition, the medication was changed to itraconazole (200 mg/day),
and latter, aiming a better CNS penetration, to fluconazole (400 mg/day). The
azoles were administered for 11 weeks, and during this period the fever resolved
and neurological symptoms stabilized. No significant collateral reaction was
observed and three months after treatment the xenodiagnosis was negative and
the titer of indirect hemagglutination test was 1:16 (Solari et al. 1993). Following
the same methodology described for the treatment with allopurinol, Apt et al.
(1998) treated 135 chronic patients with itraconazole (6 mg/kg/day for 120 days)
observing parasitological cure and normalization of ECG in 36.5% of the treated
patients but new abnormalities of the ECG appeared in 48.2% after treatment.
As in the case of allopurinol these azoles must be further investigated in a
well-designed protocol for treatment of chagasic patients.
The azoles fluconazole (a-(2,4-difluorophenyl)-a-(1H-1,2,4,-triazol-1-ylmethyl)-1H-1,2,4-triazol-1-ethanol,
Fig. 1e) and itraconazole (cis-4[4-4-4[[2-(2-4-dichlorophenyl)-2-(1H-1,2,4,triazol-1-methyl)-1,3-dioxolan-4-yl]-1
-piperazinyl]phenyl]-2,4-dihydro-2-(1-methyl-propyl)-3H -1,2,4-triazol-3-one,
Fig. 1f) have been previously assayed in
experimental animals, and their mechanism of action against T. cruzi
involve interference on ergosterol synthesis (reviewed in De Castro 1993). A
more recent study showed potent effect of the D(+) isomer of fluconazole –
compound D0870 – in both acute and chronic mice models, with 30-50 times
higher activity than ketoconazole and Nif and leading to 60-70% of parasitological
cure (Urbina et al. 1996). A formulation of D0870 as loaded nanospheres administered
by intravenous route to mice, showed also a significant cure rate (Molina et
al. 2001). It is important to stress that this compound, based on several cure
parameters was also active in a chronic phase model. We believe that the pharmacokinetics
of D0870 is now being investigated.
DEVELOPMENT
OF NEW DRUGS
Development of anti-parasite chemotherapy
could emerge from screening of synthetic or natural libraries, of compounds
with structural similarities, with a drug with recognized activity, of assays
with agents already approved for other diseases or through the determination
of a specific target, identified in a key metabolic pathway. Although several
putative targets have been presented, there is a need for their validation.
The criteria for such validation was discussed by Wang (1997), who suggested
that preliminary verifications can be indicated by in vitro activity of an inhibitor
of the putative target, but before a major effort is directed to the design
of specific inhibitors, three approaches should be used: (i) correlation of
the target inhibition and anti-parasite activity among a series of drug derivatives;
(ii) the comparison of the target between drug-sensitive and drug-resistant
parasites; (iii) the knock out of the gene encoding such target in the parasite.
In this paper Wang (1997) also pointed out the inherent difficulties of such
approaches of target validation.
Promising targets
Recent developments in the study
of the basic biochemistry of T. cruzi have allowed the identification
of novel targets for chemotherapy that include sterol metabolism, enzymes such
as trypanothione reductase, cystein proteinase, hypoxanthine-guanine phosphoribosyltrans-ferase,
glyceraldehyde-3-phosphate dehydrogenase, DNA topoisomerases, dihydrofolate
reductase and far-nesylpyrophosphate synthase (reviewed in DoCampo 2001 and
in Rodriguez 2001).
Sterol synthesis
The knowledge about sterol synthesis
on fungi opened the possibility of interference in this pathway, leading several
pharmaceutical companies to develop drugs for the treatment of different types
of superficial mycosis and systemic fungal infections. Since the main sterol
of T. cruzi is ergosterol, an intensive and fruitful investigation about
the potential effect of inhibitors of this sterol, especially by the group of
Urbina in Venezuela. Parallel to clinical studies with azole derivatives, the
trypanocidal activity and mechanism of action of new compounds is under intensive
investigation.
The triazole posaconazole (SCH56592,
Schering-Plouch), inhibited epimastigote proliferation and ergosterol synthesis
at levels 30 to 100 times higher than ketoconazole and D0870. In experimental
infections, this compound led to a cure rate of 50% in animals infected with
strains resistant to Nif, Bz and ketoconazole (Molina et al. 2000). Another
triazole derivative UR-9825 was very active against epimastigotes and intracellular
amastigotes. At the minimum inhibitory concentration for epimastigotes occurred
also depletion of 4,14-desmethyl endogenous sterols, such as ergosterol, and
their replacement by methylated sterols, indicating inhibition of C14-alpha
demethylase, as previously reported for other azoles. This drug induced also
alteration in the phospholipid profile of the parasite (Urbina et al. 2000).
The induction of resistance of T.
cruzi to azoles, such as fluconazole, and also the cross resistance between
ketoconazole, miconazole and itraconazole, observed in in vitro experiments
point to difficulties in the use of such compounds as chemotherapeutic agents
(Buckner et al. 1998). In a subsequent work, Buckner et al. (2001) reported
the development of inhibitors of a key enzyme in sterol biosynthesis, oxidosqualene
cyclase, which converts 2,3-oxidosqualene to lanosterol. The lead compound,
N-(4E,8E)-5,9, 13-trimethyl-4,8, 12-tetradecatrien-1-ylpyridinium, was shown
to cause an accumulation of oxidosqualene and decreased production of lanosterol
and ergosterol in T. cruzi. This compound and 27 related derivatives
were tested against T. cruzi, and 12 of them were highly active against
trypomastigotes.
Trypanothione reductase
Trypanosomatids present trypanothione
(N1,N8-bis(glutationyl)spermidine) and of specific enzymes
for this cofactor, trypanothione reductase (TR) and trypanothione oxidase (reviewed
in Fairlamb & Cerami 1992). TR is an NADPH-dependent flavoprotein that maintains
trypanothione in its reduced form and able to be oxidized by trypanothione oxidase,
leading to reduction of free radicals levels and contributing to the maintenance
of an intracellular reducing environment. TR has been used as a target for rational
drug design against trypanosomiasis and leishmaniasis in a number of laboratories,
since this enzyme and the mammalian counterpart (gluthatione peroxidase/glutathione
reductase system) differ on the substrate specificity (reviewed in Augustyns
et al. 2001). The determination of the structure of the active center of TR
(Krauth-Siegel et al. 1987) allowed the search of inhibitors of this enzyme,
being assayed different classes of compounds. In most cases the studies analyzed
the effect of a putative inhibitor on the purified enzyme, and depending on
the results obtained, new compounds based on molecular modeling were developed.
A first group of inhibitors reported were the so-called "subversive substrates",
due to the futile-cycling of TR induced by redox-damaging drugs, such as nitrofurans,
and naphtoquinones (Henderson et al. 1988, Salmon-Chemin et al. 2001). Subsequently
the structure of tricyclic neuroleptic showed to be a promising backbone class
of TR inhibitors, and based on computational design techniques several tricylcic
compounds were investigated (Chan et al. 1998, Gutierrez-Correa et al. 2001).
Some compounds of the series of 2-amino diphenylsulfides, that have lower neuroleptic
activity than phenothiazines, were potent inhibitors of TR (Girault et al. 1998).
Polyamine derivatives (Bonnet et al. 1997, Li et al. 2001), bisbenzylisoquinoline
alkaloids (Fournet et al. 1998) and platinum II complexes (Bonse et al. 2000)
were also studied in their capacity of inhibiting TR of T. cruzi.
Cystein protease
Cruzipain, also known as cruzain
or GP57/51, is a member of the papain C1 family of cystein proteinases (CPs).
The T. cruzi enzyme consists of a catalytic moiety with high homology
to cathepsins S and L, and is absent in all other C1 families described so far
(reviewed in Cazzulo et al. 2001). Irreversible inhibitors of cruzipain, such
as several peptidyl diazomethylketones, peptidyl fluoro-methylketones and peptidyl
vinyl sulphones interfered with the in vitro intracellular cycle of T. cruzi,
killing the parasite (reviewed in McKerrow 1999).
The treatment of acutely infected
mice with the vinyl sulphone N-piperazine-Phe-hPhe-vinyl sulphone phenyl led
to the absence of myocardial lesions, lymphocyte infiltration and intracellular
amastigote clusters. This drug kills T. cruzi by inducing an accumulation
of unprocessed cruzipain in the Golgi cisternae, interfering with the secretory
pathway (Engel et al. 1998a,b). Cruzipain exposed to biotin-labelled peptidyl
diazomethane inhibitors with a spacer arm showed a stronger reaction than the
counterparts without such spacer, probably due to differences in the topologies
of the binding sites of proteinases, differences that could be exploited to
improve specificity against trypanosomal CP (Lalmanach et al. 1996). Roush et
al. (2000) substituting the L-leucine residue of the natural peptidylepoxysuccinate
E-64, a selective irreversible inhibitor of CP, by a D-threonine obtained
a derivative with much higher activity against cruzipain than against bovine
cathepsin B. Yong et al. (2000) commented that a possible limitation of CP as
a target would be the emergence of parasite populations developing resistance
to inhibitors. These authors reported a phenotypically stable T. cruzi
cell line (R-Dm28) that displays increased resistance to Z-(SBz)Cys-Phe-CHN2,
an irreversible cysteine proteinase inhibitor, which preferentially inactivates
cathepsin L-like enzymes.
Hypoxanthine-guanine phosphoribosyltransferase
Trypanosomatids must rely upon the
salvage of exogenous purines for nucleotide synthesis, while in mammals these
nucleotides are synthesized both de novo and salvaged from recycled purine
bases. These protozoa convert purine bases to ribonucleotides, by the single
enzyme HGPRT. This enzyme can also initiate in these parasites the metabolism
of certain cytotoxic purine base analogs, such as allopurinol. This implies
that either inhibitors or substrates of HGPRT have the potential of being effective
and selective chemotherapeutic agents. The hgprt genes from T. cruzi
and other pathogenic trypanosomatids have been cloned, sequenced and overexpressed
in Escherichia coli, and the recombinant proteins have all been purified
and characterized (reviewed in Ullman & Carter 1997).
The purine analogs 3'-deoxyinosine,
3'-deoxyadenosine and allopurinol inhibited the proliferation of amastigotes
in HeLa cells, being the latter the most active. Among the pyrimidine analogs,
3'-azido-3'-deoxythymidine showed high activity against T. cruzi (Nakajima-Shimada
et al. 1996). Purine analogs were assayed for their interaction with the HGPRTs
from T. cruzi and man and some of them showed affinity for the trypanosomal
enzyme (Eakin et al. 1997). A structure-based docking method identified 22 potential
inhibitors of the enzyme. Three compounds (2,4,7-trinitro-9-fluorenyl-idenemalononitrite,
3-(2-fluorophenyl)-5-(phenoxy)-1,2,4-triazolo (4,3-C)-quinazoline and 3,5-diphenyl-4´-methyl-2-nitrobiphenyl)
were effective against intracellular amastigotes and one [6-(2,2-dichloro-aceta-mido)chrysene]
was a potent inhibitor of the trypanosomal HPRT (Freymann et al. 2000).
DNA topoisomerases
DNA topoisomerases II are enzymes
that alter the topology of DNA and in kinetoplastids have been the focus of
considerable study in the areas of molecular and cellular biology and also experimental
chemotherapy. The gene encoding T. cruzi type II topoisomerase was isolated
and the comparison with the amino acid sequence of the corresponding enzymes
of T. brucei and Crithidia fasciculata showed a high degree of
conservation (Fragoso & Goldenberg 1992). The enzyme is expressed in epimastigotes
but not in trypomastigotes, although both forms of the parasite present the
mRNA encoding the enzyme and is localized exclusively in the nucleus of the
parasite (Fragoso et al. 1998).
Several inhibitors of bacterial DNA
topoisomerase II showed activity against T. cruzi, inhibiting both proliferation
and differentiation processes, and causing damage to kinetoplast and/or the
nucleus of epimastigotes (Kerschmann et al. 1989, Gonzales-Perdomo et al. 1990),
suggesting that both organelles could be the targets of the drugs. Camptothecin,
inhibitor of eukaryotic DNA topoisomerase I, induced cleavage of nuclear and
mitochondrial DNA in T. cruzi (Bodley & Shapiro 1995).
Dihydrofolate reductase
Dihydrofolate reductase (DHFR) and
thymidylate synthetase exist as a bifunctional protein in different species
of protozoa. This enzyme has successfully been used as a drug target in chemotherapy
of cancer, malaria and bacterial infections. The gene coding for the
DHFR domain from T. cruzi was cloned and expressed (Reche et al. 1996).
Zucotto et al. (1998) described the modelling of T. cruzi's DHFR based
on the crystal structure of Leishmania major enzyme. From methotrexate,
inhibitor of the human enzyme, among several derivatives synthesized, some of
them showed a greater selectivity for the parasite enzyme than for the human
counterpart (Zuccotto et al. 1999). In the same line, Chowdhury et al. (2001)
designed and synthesized novel inhibitors of DHFR of trypanosomatids, however
the compounds showed weak activity against both the enzyme and intracellular
amastigotes of T. cruzi.
Glyceraldehyde-3-phosphate dehydrogenase
Since intracellular amastigotes possibly
derive its energy from glycolysis, inhibition of glycolytic enzymes such as
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) may be a novel approach for
the development of anti-T. cruzi drugs. The structure of GAPDH from glycosomes
was reported and comparison with that of the mammalian counterpart led to the
group of Oliva to consider the possibility of development of specific inhibitors
of the parasite enzyme (Souza et al. 1998). In a subsequent work the isolation
of flavonoids from the fruits of Neoraptua magnifica led to the compound
3',4',5',5,7-pentamethoxy-flavone that showed the highest activity over flavones
and pyrano chalcones against the GAPDH of the parasite (Tomazela et al. 2000).
Crystal structure of trypanosomatids and human GAPDHs provided details about
the interaction of adenosyl moiety of NAD+ with proteins. Although
adenosine is a very poor inhibitor, addition of substituents to the 2' position
of ribose and the N6-position of adenosine led to a series of disubstituted
nucleosides, and [N6-(1-naphthalenemethyl)-2'-(3-chlorobenzamido) adenosine]
inhibited the proliferation of amastigotes without effect on the corresponding
human enzyme (Bressi et al. 2001).
Farnesylpyrophosphate synthase
In pathogenic protozoa the pathway
responsible for the synthesis of a variety of sterols and polyisoprenoids involves
the enzyme farnesylpyrophosphate synthase, leading to the formation of farnesylpyrophosphate
that marks the branching point of these synthetic routes. A gene encoding the
farnesylpyrophosphate synthase of T. cruzi (TcFPPS) was cloned and sequenced
and the enzyme was inhibited by the nitrogen-containing bisphosphonates, such
as pamidronate and risedronate, but was less sensitive to the non-nitrogen-containing
bisphosphonate etidronate, which does not affect parasite growth (Montalvetti
et al. 2001). Pamidronate caused a decrease in the parasitemia of T. cruzi-infected
mice and inhibited the in vitro intracellular replication of amastigotes (Urbina
et al. 1999). Risedronate inhibited the proliferation of epimastigotes and sterol
biosynthesis at a pre-squalene level and based on sterols analysis in treated
parasites Martin et al. (2001) associated these results with the inhibition
of farnesylpyrophosphate synthase. The effect of a series of bisphosphonates
derived from fatty acids were assayed against T. cruzi and some of these
drugs were potent inhibitors of the proliferation of intracellular amastigotes,
but all of them were devoid of activity against epimastigotes (Szajnman et al.
2000). The selective action of nitrogen-containing bisphophonates against T.
cruzi in comparison to mammalian cells could result from the preferential
drug accumulation in parasite acidocalcisomes, acidic organelles rich in calcium,
pyrophosphate, magnesium, sodium, zinc and polyphosphates (reviewed in DoCampo
& Moreno 2001).
Experimentally assayed drugs after
1992/1993
The present review offers a survey
of the available literature about new classes of compounds and also new derivatives
from compounds previously assayed in the search for new drugs against T.
cruzi. Recently other reviews have also been published, most of them exploring
selected groups of compounds or inhibitors for T. cruzi targets (Rodriguez
& Gros 1995, Urbina 1999, DoCampo 2001, Rodriguez 2001). As we have already
mentioned with the present review, together with those of Brener (1979) and
De Castro (1993), we intend to give the reader a general view of the experimental
studies in the area of the chemotherapy of Chagas disease.
Synthetic drugs
Thiadiazine derivatives -
In assays with epimastigotes, most of the 1,3,5-thiadiazine-2-thione derivatives
were more active than Nif, while among 1,2,6-thiadiazin-3,5-dione 1,1-dioxides;
although active against the parasite, most of them were also toxic to mammalian
cells (Ochoa et al. 1999, Di Maio et al. 1999). Among the less cytotoxic derivatives
of the second series, one compound showed activity against intracellular amastigotes
similar to the standard drug (Muelas et al. 2001).
1,2,5-oxadiazole N-oxide derivatives
- A series of 1,2,5-oxadiazole N-oxide, benzo[1,2-c]1,2,5-oxadiazole N-oxide,
and of quinoxaline di-N-oxide derivatives were synthesized and the activity
against epimastigotes was associated with N-oxide radical formation (Cerecetto
et al. 1999).
1,4-dihydropyridines - Among
nitro-aryl-1,4-dihydropyridines, nicardipine, isradipine and lacidipine inhibited
epimastigote proliferation and oxygen uptake in intact parasite, and the first
compound showed also a similar effect in mitochondria in situ (Nunes-Vergara
et al. 1997). For a series of 3-chloro-phenyl-1,4-dihydropyridine derivatives
a positive correlation between trypanocidal effect and easiness of oxidation
of the dihydripyridine ring was found (Maya et al. 2000).
Acridine derivatives - Since
the 80s several acridine and acridinone derivatives have been presented a correlation
of activity against epimastigotes and DNA binding (reviewed in De Castro 1993).
Among a series of bis(9-amino-6-chloro-2-methoxyacridines), a bisacridine
containing piperazine as central amine showed co-localization with kDNA of epimastigotes
(Girault et al. 2000), reinforcing that their activity is associated with DNA
interaction. Several 9-thioalkylacridines were active against T. cruzi
(Bsiri et al. 1996). 9-Amino and 9-thioacridines have been reported to inhibit
the enzyme TR (Bonse et al. 2000).
Nitroimidazoles and nitrofurans
derivatives - Several nitroimidazoles such as megazol (CL 64,855), MK-436
and fexinidazole presented high activity in infected animals (reviewed in De
Castro 1993). The current investigation about the use of nitroimidazoles for
therapy of African trypanosomiasis, especially megazol (Barrett et al. 2000),
renewed the interest in the area of Chagas disease. The coupling of 5-chloro-4-nitro-1-methylimidazole
with heterocycles led to the synthesis of two compounds with activity against
trypomastigotes (Boechat et al. 2001). More recently Cerecetto et al. (1999)
synthesized nitrofurazones (5-nitro-2-furaldehyde semicarbazones) and tyophenes
(5-nitrothiophene-2-carboxaldehyde) in which N4-semicarbazone moiety
was replaced by different of amines, aiming to mimic the spermidine part of
trypanothione. These compounds presented lower activity against epimastigotes
than the parent compound, while some nitrofurazones bearing N4 other
substituents produced complete survival in infected mice (Cerecetto et al. 2000).
Phenothiazines - Before the
period mentioned in this review several groups investigated the effect of phenothiazines,
tricyclic compounds employed clinically as antidepressants (reviewed in De Castro
1993). More recently it was reported that this class inhibited the enzyme TR.
Promethazine and thioridazine assayed in vivo decreased parasitemia levels and
mortality (Paglini-Oliva et al. 1998). When mice infected with low innoculum
of T. cruzi and treated with thioridazine were checked 135 days post-infection,
the heart histology and density of cardiac b-receptors
were similar to those of uninfected, untreated controls (Rivarola et al. 1999),
suggesting to the authors that this drug could prevent the evolution to the
chronic phase of the infection.
Metal chelating agents and metallic
complexes - Several chelating agents and derivatives were active against
epimastigotes; being proposed that they act by interfering with the essential
metabolism of iron, copper, or zinc (Rodrigues et al. 1995). Several Fe3+
chelating agents showed activity against epimastigotes, that could be decrease
in the presence of iron (Jones et al. 1993, Singh et al. 1997), and tetraethyl
derivative of aminothiol was active against trypomastigotes (Deharo et al. 2000).
The metal chelator sodium diethylamine-N-carbodithioate showed, in relation
to Bz a similar activity against epimastigotes and intracellular amastigotes
but lower activity against trypomastigotes (Lane et al. 1996). Another chelator,
1,10-phenanthroline inhibited epimastigote proliferation and led to electron-dense
deposits in the kinetoplast, mitochondrion, and endoplasmic reticulum, identified
as containing predominantly calcium and suggesting to the authors the involvement
of disruption of calcium homeostasis in the trypanocidal activity (Lane et al.
1998).
Among several osmium(III) complexed
with carbamates and different metals complexed with 1,2,4]triazolo [1,5a]pyrimidines
some compounds were active against epimastigotes (Castilla et al. 1996, Luque
et al. 2000).
Propene-1-amine derivatives -
A series of 3-(4'-bromo-[1,1'-biphenyl]-4-yl)-3-(4-X-phenyl)-
N,N-dimethyl-2-propen-1-amine derivatives were active against the three
forms of T. cruzi (De Conti et al. 1996, Oliveira et al. 1999). The two
compounds – the unsubstituted (X = H) and the bromine (X =
Br) analogs – with highest activity against bloodstream forms, and lowest
toxicity to mammalian cells, were assayed in vivo. The bromo-derivative displayed
a strong suppressive effect on the parasitemia, and led to the survival of all
the treated mice, whereas its unsubstituted analog was ineffective under the
same conditions (Pereira et al. 1998).
Aminoquinoline derivatives -
Quinolines have been assayed as potential drugs for Chagas disease since the
50s (reviewed in Brener 1979). Against epimastigotes the action of primaquine
involves the formation of free radicals and this drug presented synergistic
effect with ketoconazole decreasing the parasitemia of experimentally infected
mice (reviewed in De Castro 1993). More recently, dipeptide derivatives of this
aminoquinoline were synthesized as prodrugs, and inhibited the infection of
LLC-MK2 cells with T. cruzi (Chung et al. 1997). Among 77 primaquine
analogues, one of them reduced the parasitemia in mice in levels 14 and 4 times
higher than Nif and primaquine, respectively (Kinnamon et al. 1996). In the
same model, among 40 8-aminoquinolines, non-related to primaquine, 6 were more
active than Nif, and for one of them, the activity was 13 times higher than
this standard drug (Kinnamon et al. 1997).
Dinitroanilines - These compounds
are microtubule-disrupting herbicides and one of the most studied is trifluralin.
This compound was active against epimastigotes, trypomastigotes, and axenic
and intracellular amastigotes. In trypomastigotes, trifluralin led to alterations
at the surface analyzed by transmission and scanning electron microscopy, with
membrane waving not associated with sub-pellicular microtubules. Treated epimastigotes
showed alterations of shape, and some parasites presented two or three flagella
and kinetoplasts, suggesting interruption of the cytokinesis process. Trifluralin
also inhibited endocytosis in epimastigotes, monitored by complexes of gold
with bovine serum albumine (Dantas et al. 1998, Dantas 2000). This dinitroaniline
presented also in vivo effect in mice model (Zaidenberg et al. 1999) and inhibited
the differentiation of epimastigotes to trypomastigotes (Bogitsh et al. 1999).
Recently we reviewed the effect of
dinitroanilines on pathogenic protozoa and observed that Leishmania spp.
and Trypanosoma brucei were more susceptible to trifluralin than T.
cruzi and also compared sequences of tubulins of susceptible organisms (plants
and trypanosomatids) and resistant (mammals) (Traub-Cseko et al. 2001).
Lysophospholipid analogs -
These compounds are under clinical studies for cancer chemotherapy. The most
studied derivatives are the alkylglycerophosphocholine edelfosine, a thioether
substituted phosphatidylcholine analog ilmofosine and the alkylphosphocholine
hexadecylphosphocholine (mitelfosine) (reviewed in Wieder et al. 1999). Clinical
trials with mitelfosine for the treatment of Indian visceral leishmaniasis gave
high percentage of cure including in cases resistance antimony therapy (Sundar
et al. 2000). These LPAs were active against epimastigotes, intracellular amastigotes
and trypomastigotes and led to damage of the flagellar membrane of epimastigote,
and edelfosine inhibited the in vitro metacyclogenesis process (Santa-Rita et
al. 2000). Previous in vivo experiments showed that LPAs had only a suppressive
effect on the parasitaemia of T. cruzi-infected mice (Croft et al. 1996).
It was found that LPAs are potent inhibitors of phosphatidylcholine synthesis
in epimastigotes, and that ergosterol and its 24-ethyl analog were replaced
by its D22-saturarted analogs, indicating
inhibition of sterol C-22 desaturase (Lira et al. 2001).
Drugs derived from natural sources
Natural products account for about
half of the pharmaceuticals in use today, but there has been a shift away from
their use with the increasing predominance of molecular approaches to drug discovery
(Clark 1996). However, in the last decades occurred a movement named by some
as "back to Nature", with the revival of phytotherapy, needing a multidisciplinary
group with expertise in botany, chemistry, biochemistry, molecular and cellular
biology, and pharmacology. This trend was accentuated by the success obtained,
for example, by taxol in cancer chemotherapy and artemisinin for malaria. A
complementary approach is the development of synthetic analogs of the lead natural
compound, aiming the improvement of properties such as pharmacokinetics, compatibilities
and stability. Such intensification in the search of drugs from natural sources
was also observed in the area of trypanosomiasis and leishmaniasis. In the literature
several recent reports deal with the investigation of trypanocidal activity
of a wide variety of crude natural extracts, especially vegetal ones, compounds
isolated and semi-synthetic analogs.
Alkaloids - A great number
of alkaloids have been assayed against T. cruzi. The activity of a series
of alkaloids against epimastigotes was associated to the inhibition of cell
respiration, and the most active compound was apomorphine (Morello et al. 1994).
Using epimastigotes of Nif-resistant, the activity of b-carboline
alkaloids was also associated with respiratory chain (Rivas et al. 1999). Several
glycoalkaloids were tested against epimastigotes, bloodstream and metacyclic
trypomastigotes, and a-chaconine and a-solamargine showed higher activity than
ketoconazole (Chataing et al. 1998). Another group studied was bisbenzylisoquinoline
alkaloids and daphnoline and cepharanthine were active against the parasite
and inhibited enzyme TR (Fournet et al. 1998). In acute infections, dapholine
led to a significant decrease in parasitemia and increase in cure rate in comparison
with Bz-treated mice, and in chronic infections, in 70% of the treated mice
no parasite was detected (Fournet et al. 2000). Five new bisbenzylisoquinoline
derivatives were isolated from the stem bark of Guatteria boliviana and
funiferine, antioquine and guatteboline were active against trypomastigotes
(Mahiou et al. 2000). The alkaloid anti-microtubule agents vinblastine and vincristine
obtained from Vinca rosea interfere with the proliferation of
epimastigotes, inhibiting both nuclear division and cytokinesis, leading to
giant cells with multiple nuclei, kinetoplasts and flagella (Grellier et al.
1999).
Taxoids - Taxol obtained from
the bark of Taxus brevifolia is an anti-microtubule drug. This compound
and synthetic derivatives are employed in cancer chemotherapy. Taxol was active
against epimastigotes and trypo-mastigotes leading to significant alterations
in the morphology of the parasites. Taxol also inhibited the endocytosis of
proteins by epimastigotes (Dantas 2000).
Stilbenoids and derivatives -
Among natural dihydrostilbenoid isonotholaenic acid and several simple derivatives
some compounds showed activity similar to Bz against epimastigotes and others
were more active than crystal violet against trypomastigotes (Olmo et al. 2001).
Snake venom - Venom from Cerastes
cerastes and Naja haje inhibited the proliferation of epimastigotes
of T. cruzi at levels similar to Bz, and that of C. cerastes was
also active against trypomastigotes (Fernandez-Gomez et al. 1994).
Gangliosides Treatment with
gangliosides of mice during acute infection promoted survival and clearance
of parasites from the bloodstream and organs, and it was suggested that the
effect of gangliosides could be due to interference of parasite penetration
into the host cells due to inhibition of phospholipase A2 (Lujan
et al. 1993), but since the compounds had no direct effect on the parasite,
Bronia et al. (1999) consider that the in vivo effect could be due to modulation
of the host immune system.
Juvenile hormone inhibitors and
analogs - The juvenile hormone fenoxycarb and methoprene inhibited the proliferation
of epimastigotes (Stoka et al. 1995). Fenoxycarb and analogs were synthesized,
and assays with epimastigotes suggested that an allyl ether moiety bonded at
the polar extreme is important for the trypanocidal effect (Cinque et al. 1998).
Some analogs were also active against bloodstream forms and reduced the parasitemia
and mortality levels in relation to untreated controls (Fichera et al. 1995).
4-Phenoxyphenoxyethyl thiocyanate was very active against epimastigote and accumulation
of low molecular weight metabolites from mevalonate to squalene was observed
(Szajnman et al. 2000), leading the authors to suggest that the effect is associated
with interference in the synthesis of ergosterol. Also sulfur-containing derivatives
showed high activity against epimastigotes and amastigotes of T. cruzi
(Rodriguez et al. 2000).
Propolis - It is a natural
resin produced by honey bees has been used in folk medicine, since it displays
strong anti-microbial activity, associated mainly with flavonoids and derivatives
of hydroxycinamic acid (reviewed in De Castro 2001). Ethanolic extract prepared
from a North American commercial sample were active in vitro against epimastigotes,
trypomastigotes and intracellular amastigotes (Higashi & De Castro 1994).
However, in vivo experiments using different propolis formulations, showed no
effect on the course of acute infection (De Castro & Higashi 1995). A Bulgarian
sample has been assayed against trypomastigotes, and the ethanolic and acetonic
extracts showed activity, respectively, similar and two times higher than crystal
violet. The chemical composition of both extracts showed a high content of flavonoids,
with differences in relative individual concentrations (De Castro et al. 2001).
Recently Brazilian propolis became a subject of increasing scientific and commercial
interest, and due to significant differences between the composition samples
form tropical and temperate zones (reviewed in Bankova et al. 2000), we isolated
from a sample collected in the State of Paraná (Brazil) four derivatives
of hydroxycinnamic acid than when assayed against trypomastigotes showed activity
bellow that of crystal violet (Marcucci et al. 2001).
Naphthoquinones and synthetic
derivatives - Quinones are present in different families of plants and in
some of them have been used in folk medicine for the treatment of diseases,
especially cancer. Among bioactive natural naphthoquinones, we found lapachol,
isolated from the heartwood of Tabebuia ("ipês") and also a-
and b-lapachone obtained as contaminants in the process
of lapachol isolation. Recently, lapachol derivatives were assayed against trypomastigotes
and the triacetoxy derivative of reduced lapachol showed relevant trypanocidal
activity (Santos et al. 2001). Due to the increasing number of reports about
the activity of b-lapachone against a wide variety
of tumor cells, by inhibition of topoisomerases and induction of apoptosis,
Dubin et al. (2001) pointed out the potential of clinical applications of this
naphthoquinone. Previous studies with
b-lapachone showed activity against epimastigotes that was associated
to generation of free radicals and inhibition of nucleic acids and protein synthesis
(reviewed in De Moura et al. 2001). In a study of naphthoquinones isolated from
Tabebuia, we synthesized 50 heterocyclic derivatives. The overall analysis of
the structures indicated the tendency of trypanocidal activity in compounds
with an imidazole or oxazole ring linked to a naphthopyrane structure (Pinto
et al. 1997, Neves-Pinto et al. 2000). Among these compounds two naphthoimidazoles
showed activities against trypomastigotes was 14.5 and 34.8 times higher than
the standard crystal violet and both have an aromatic moiety linked to the imidazole
ring (reviewed in De Moura et al. 2001).
Other groups of quinones isolated
from natural products were also assayed against T. cruzi: (a)
the trihydroxylated anthraquinone purpurin, obtained from the roots of Rubia
tinctorum (Rubiaceae), showed an activity against trypomastigotes 1.5 times
higher than crystal violet (De Castro et al. 1994); (b) among 1,4-naphthoquinones
isolated from Calceolaria sessilis, 2,3,3-trimethyl-2-3-dihydronaphtho[2,3-b]furan-4,9-quinone
showed against epimastigote and tumor lineages a high cytotoxicity and a temporary
increase of oxygen consumption (Morello et al. 1995); (c) the polyprenylated
benzoquinone 7-epiclusianone, isolated from Rheedia gardneriana (Clusiacease)
was active in vitro against trypomastigote, but showed no effect on experimentally
infected (Alves et al. 1999).
Crude plant extracts and components
– (i) The sesquiterpene lactone dehydroleucodine isolated from Artemisia
douglasssiana was active against epimastigotes, that presented pycnotic
nucleus and a decreased number of ribosomes (Brengio et al. 2000); (ii) cryptofolione
isolated from the fruits of Cryptocarya alba was active against trypomastigotes,
however showed moderate effect for both amastigotes and macrophages, indicating
little selectivity for T. cruzi (Schmeda-Hirschmann et al. 2001); (iii)
from the seeds of Annona glauca (Annonaceae) nine acetogenins were isolated,
and among them glaucanisin, squamocin, annonacin A and annonacin showed activity
against trypomastigotes (Waechter et al. 1998); (iv) megalomicin is a macrolide
antibiotic produced by Micromonospora megalomicea, that inhibits vesicular
transport in Golgi apparatus, resulting in the undersialylation of cellular
proteins in mammalian cells. This compound was active against epimastigotes
and intracellular amastigotes at concentrations bellow those that interfere
with the mammalian organelle (Bonay et al. 1998); (v) from the leaves of Zanthoxyllum
naranjillo (Rutaceae), the lignan, methylpluviatolide was highly
effective in vitro and in vivo (Bastos et al. 1999); (vi) Terpenes: (a)
among diterpenoids isolated from Azorella compacta. (Llareta) azorellanol
and mulin-11,13-dien-20-oico acid were active against intracellular amastigotes
and the cytotoxicity to mammalian cells was lower than that of Nif (Neira et
al. 1998); (b) from the aerial parts of Wedelia paludosa (Asteraceae),
the isolated diterpenes ent-kaur-16-en-19-oic acid, ent-kaur-9(11),16(17)-dien-19-oic
acid and 3 alpha-angeloiloxy-ent-kaur-16-en-19-oic acid showed activity against
trypomastigotes (Batista et al. 1999); (c) in a study in Bolivia, 14
plants used in folk medicine to treat cutaneous leishmaniasis, extracts from
53 medicinal plants used for other diseases and different parts from 43 plants
were screened and some of these material showed activity against epimastigotes
of different strains of T. cruzi (Fournet et al. 1994); (d) extracts
used in popular medicine in Guatemala from several plants such as Neurolaena
lobata (Asteraceae), Tridax procumbens (Asteraceae), Petiveria
alliacea (Phytolaccaceae) and Byrsonima crassifolia (Malpighiaceae)
showed high activity against trypomastigotes. N. lobata and Solanum
americanum showed in vitro and in vivo trypanocidal activity (Caceres et
al. 1998). In a subsequent work, extracts, fractions and isolated sesquiterpene
lactones and germacranolides from N. lobata showed high activity against
epimastigotes (Berger et al. 2001); (e) among 79 extracts from plants
of the families Asteraceae, Araceae, Moraceae, Solanaceae, Rhamnaceae, Zingiberaceae,
Leguminosae and Sapotaceae, nine of then were active against epimastigotes (Muelas-Serrano
et al. 2000); (f) from 32 crude plant extracts of nine species of Rutaceae
eight of them showed significant activity trypomastigote. Fractionation of the
active extracts provided 25 fractions, and the two fractions more active were
obtained from the leaves of Almeidea coerulea and from Conchocarpus
inopinatus (Mafezoli et al. 2000).
Prophylatic drugs
Although recent advances in vector
control in the Southern Cone countries, by initiative of the Pan American Health
Organization (PAHO) and World Health Organization (WHO), have decreased the
incidence of new infections (Schofield & Dias 1999), we are still challenged
by two critical problems: the treatment of chronic cases of the disease and
the high level of acute cases in some Latin American countries, such as Bolivia
and Mexico, where the incidence of infection in some regions reaches levels
above 80% of the population (Medrano-Mercado et al. 1996). In endemic areas
the transfusional transmission of Chagas disease, due mainly to urbanization
and migration processes, represents a great threat (Wendel & Dias 1992).
For example, in 1990 in the USA, 7 million persons emigrated from countries
in which Chagas disease is endemic (Schmunis, 1994, 2000) a preoccupying fact
for the health authorities of non-endemic countries.
There is a recommendation from the
WHO (1984) for the use of crystal violet in hemotherapic centers in endemic
areas to eliminate the parasite in the blood used for transfusion (Nussenzweig
et al. 1953). This dye presents no substantial side effect, although there are
reports about blood micro-agglutination and potential mutagenicity (Thomas &
McPhee 1984). Its main disadvantage is the bluish color that it confers to blood
and tissues, which is not well accepted by the population. The development of
trypanocidal drugs also involves the search for alternatives for crystal violet,
when the assayed were performed with trypomastigotes in the presence of blood.
These studies are included in the above item, but it must be kept in mind that
when looking for an alternative to crystal violet its is fundamental to perform
the assays with bloodstream forms in the presence of blood and at 4°C. The
presence o blood is essential, since several trypanocidal compounds that are
inactivated by blood components, such as gossypol (Rovai et al. 1990) and naphthoquinones
and their heterocyclic derivatives (Lopes et al. 1978, Pinto et al. 1997). Another
problem associated with drugs for prophylaxis of blood is, besides eventual
toxicity, the poor solubility in aqueous medium and binding to plasma protein.
CONCLUDING
REMARKS
Nowadays infectious diseases are
still an important cause of mortality and morbidity and the rising incidence
of emerging or re-emerging diseases can be explained, at least partly, by the
deterioration of health care systems and diverse socio-economic and ecological
disorders. The cost of investments and the lack of market potential and market
security in developing countries have dampened interest in developing drugs
for tropical diseases. Among the 1061 new drugs developed from 1975 to 1994,
less than 2.7% concern tropical diseases (Trouiller et al. 2000).
In relation to Chagas disease, great
advances are being made in parts of South America to control the transmission
by insect vectors or blood transfusion, but more effective chemotherapy is needed
for the millions who are already infected. The demonstration of parasite in
chronic patients indicates its importance in the maintenance of Chagas disease
(reviewed in Tarleton 2001) and open the discussion about the etiologic treatment
during this phase (OPAS/OMS 1998), reinforcing the need of finding more efficient
and less toxic drugs.
In relation to experimental studies,
a dichotomy has arisen between a rational and an empirical approach for drug
development (Croft 1994). However, both are important routes to achieve a potential
drug and sometimes are even used in conjunction. For example, having in hands
a potential parasite target, the finding of an inhibitor could be searched by
empirical tests or by modeling and synthesis that could interfere with the special
regions of the target, such as the active center of an enzyme. The advent of
genomics, rapid DNA sequencing, bioinformatics, combinatorial chemistry and
automated high-throughput screenings had strength the interaction among groups
with different expertise in order to find compounds with efficacy, including
for immunosuppressed patients, without or at least with low toxicity, favorable
pharmacokinetics and permeability properties and low cost of production.
Most of the studies with designed
drugs against potential targets in T. cruzi involves inhibition tests
using purified enzymes such as TR, CP and HGPRT, and it is possible that a compound
discarded due to a negative result in this assay could be effective against
the intact parasite. The continuous research of TR as a target could lead to
an inhibitor that is a trypanocidal agent itself or that compromising the redox
defenses of the parasite act in synergy with redox-damage drugs. However since
the identification of trypanothione and although several groups are involved
in finding inhibitors of TR, no drug emerged as effective and with low toxicity
in experimentally infected animals.
After the introduction of Nif and
Bz, among the extensive list of classes of compounds with in vitro and in vivo
activity against T. cruzi, with exception of allopurinol, itraconaozle
and fluconazole, none was submitted to clinical assays. This is due is some
cases, to the absence of strong indication of their curative effect, to their
potencial toxic and/or teratogenic effect (usually assayed only in vitro). This
analysis emphasizes the need of a better choice of experimental models and standardization
of protocols. In in vitro tests, it is of fundamental importance the
use of trypomastigotes in the presence of blood both at 4°C (blood prophylaxis)
and at 37°C (treatment) and comparison with crystal violet, and the effect
against intracellular amastigotes using a suitable host cell and comparison
with Nif or Bz. Still today several drugs are screened only against epimastigotes,
although several authors have already reported different sensibilities to drugs
among different forms of the parasite. In relation to in vivo tests a standardization
of both acute and chronic phase models and of parameters to monitor the cure
are of outmost importance. Also toxicity and mutagenicity studies in animals
are needed. Otherwise we will continue to employ a lot of effort in experimental
studies without achieving a drug that could be submitted to clinical trials.
After the presentation of potential
targets in the parasite and active drugs in experimental models, which are the
compounds that could potentially be employed in clinical tests? Among them stand
out the nitroimidazoles megazol (CL-64855) (Filardi & Brener 1982) and MK-436
(Andrade et al. 1989), drugs studied before 1993, but that afterwards, at least
to our knowledge, no new papers dealing with such promising drugs appeared in
the literature. We believe that it could be important to review all the data
obtained about megazol and MK-436, and develop new in vivo protocols to monitor
their pharmacokinetics and bioavailability properties and also any potential
toxic, mutagenic and/or teratogenic effects. It is possible that the discontinuity
of such studies was due to the toxicity of the compounds or no interest of pharmaceutical
companies in their synthesis. Drugs for Chagas disease are not in the interest
of such industries and only the effort and persuasion of researchers in establishing
partnerships could change the current scenery. An interesting case is megazol,
which although previously shown by the group of Brener as an active drug even
against strains of T. cruzi resistant to Nif and Bz, was not further
investigated (Filardi & Brener 1982). However, several recent works pointed
to its usefulness for the treatment of African trypanosomiasis (Barrett et al.
2000) and it was communicated in the 4th CostB9 Meeting in Portugal (2001) that
a pharmaceutical industry will probably begin to synthesize megazol with this
objective.
What must be implemented to achieve
new effective and less toxic drugs for the treatment of Chagas disease? With
all the knowledge accumulated about the biology and biochemistry of T. cruzi
a lot of effort must be directed to the understanding of the mechanism of action
of selected compounds. Another line that is beginning to be studied is the preparation
of different formulations of drugs, which would allow its delivery to the right
places. Excellent results have been obtained in the case of formulations of
amphotericin for the clinical treatment of leishmaniasis (Sundar et al. 1998)
and also experimental Chagas disease (Yardley & Croft 1999). Also in studies
with T. cruzi, it was already been shown to increase in vitro and in
vivo effect of different drugs. For example the use of ethylcyanoacry-late nanoparticles
prepared by an emulsion polymerization process together with allopurinol or
Nif and surfactants led to higher activity against epimastigotes than the corresponding
free compound (Gonzalez-Martin et al. 1998, 2000). Also Molina et al. (2001)
incorporated inhibitors of sterol biosynthesis into long-circulating polyethy-leneglycol-polylactide
nanospheres improving the bioavailability of these poorly soluble compounds
and increasing the cure rate in experimental animals.
REFERENCES
- Alves TM, Alves R, Romanha AJ, Zani CL,
Dos Santos MH, Nagem TJ 1999. Biological activities of 7-epiclusianone.
J Nat Prod 62: 369-371.
- Andrade AL, Zicker F, de Oliveira RM, Almeida
Silva S, Luquetti A, Travassos LR, Almeida IC, De Andrade SS, de Andrade
JG, Martelli CM 1996. Randomized trial of efficacy of benznidazole in treatment
of early Trypanosoma cruzi infection. Lancet 348: 1407-1413.
-
Andrade SG, Figueira RF 1977.
Estudo experimental sobre a ação terapêutica da droga
RO 7 – 1051 na infecção por diferentes cepas do Trypanosoma
cruzi. Rev Inst Med Trop São Paulo 19: 335-341.
-
Andrade SG, Andrade ZA, Figueira
RM 1977. Estudo experimental sobre a resistência de uma cepa ao Bay
2502. Rev Inst Med Trop São Paulo 19: 124-129.
- Andrade SG, Costa Silva R, Santiago CM 1989.
Treatment of chronic experimental Trypanosoma cruzi infections in mice
with MK-436, a 2-substituted 5-nitroimidazole. Bull WHO 67: 509-514.
- Andrade SG, Figueira RM, Carvalho ML, Gorini
DF 1975. Reaction of the Trypanosoma cruzi strain to the experimental
therapeutical response to Bay 2502 (results of long term treatment). Rev
Inst Med Trop São Paulo 17: 380-399.
- Andrade SG, Freitas LAR, Peyrol S, Pimentel
AR, Sadigursky M 1991. Experimental chemotherapy of Trypanosoma cruzi
infection persistence of parasite antigens and positive sorology in parasitologically
cured mice. Bull WHO 69: 191-199.
- Andrade SG, Pimentel AR, de Souza MM, Andrade
ZA 2000. Interstitial dendritic cells of the heart harbor Trypanosoma cruzi
antigens in experimentally infected dogs: importance for the pathogenesis
of chagasic myocarditis. Am J Trop Med Hyg 63: 64-70.
- Andrade ZA 1999. Immunopathology of Chagas
disease. Mem Inst Oswaldo Cruz 94 (Suppl I): 71-80.
- Anez N, Carrasco H, Parada H, Crisante G,
Rojas A, Fuenmayor C, Gonzalez N, Percoco G, Borges R, Guevara P, Ramirez
JL 1999. Myocardial parasite persistence in chronic chagasic patients. Am
J Trop Med Hyg 60: 726-732.
- Apt W, Aguilera X, Arribada A, Perez C, Miranda
C, Sanchez G, Zulantay I, Cortes P, Rodriguez J, Juri D 1998. Treatment of
chronic Chagas disease with itraconazole and allopurinol. Am J Trop Med
Hyg 59: 133-138.
- Araújo MS, Martins-Filho OA, Pereira
ME, Brener Z 2000. A combination of benznidazole and ketoconazole enhances
efficacy of chemotherapy of experimental Chagas disease. J Antimicrob Chemother
45: 819-824.
- Augustyns K, Amssoms K, Yamani A, Rajan PK,
Haemers A 2001. Trypanothione as a target in the design of antitrypanosomal
and antileishmanial agents. Curr Pharm Des 7: 1117-1141.
- Ávila H, Sigman DS, Cohen LM, Millikan
RC, Simpson L 1991. Polymerase chain reaction amplification of Trypanosoma
cruzi kinetoplast minicircle DNA isolated from whole blood lysates: diagnosis
of chronic Chagas disease. Mol Biochem Parasitol 48: 211-222.
- Bankova VS, De Castro SL, Marcucci MC 2000.
Propolis: recent advances in research on chemistry and plant origin. Apidologie
31: 3-15.
- Bastos JK, Albuquerque S, Silva ML 1999. Evaluation
of the trypanocidal activity of lignans isolated from the leaves of Zanthoxylum
naranjillo. Planta Med 65: 541-544.
- Batista R, Chiari E, de Oliveira AB 1999.
Trypanosomicidal kaurane diterpenes from Wedelia paludosa. Planta Med 65:
283-284.
- Berger I, Passreiter CM, Caceres A, Kubelka
W 2001. Antiprotozoal activity of Neurolaena lobata. Phytother Res 15:
327-330.
- Bocca-Tourres LC 1969. La enfermedad de Chagas
en período agudo y su tratamiento con el Bay 2502. Bol Chil Parasitol
24: 24-27.
- Bock M, Gonert R, Haberkorn A 1969. Studies
with Bay 2502 on animals. Bol Chil Parasitol 24: 13-19.
- Bodley AL, Shapiro TA 1995. Molecular and
cytotoxic effects of camptothecin, a topoisomerase I inhibitor, on trypanosomes
and Leishmania. Proc Natl Acad Sci USA 92: 3726-3730.
- Boechat N, Carvalho AS, Fernandez-Ferreira
E, Soares RO, Souza AS, Gibaldi D, Bozza M, Pinto AC 2001. Novel nitroimidazoles
with trypanocidal and cell growth inhibition activities. Cytobios 105:
83-90.
- Bogitsh BJ, Middleton OL, Ribeiro-Rodrigues
R 1999. Effects of the antitubulin drug trifluralin on the proliferation and
metacyclogenesis of Trypanosoma cruzi epimastigotes. Parasitol Res
85: 475-480.
- Bonay P, Duran-Chica I, Fresno M, Alarcon
B, Alcina A 1998. Antiparasitic effects of the intra-Golgi transport inhibitor
megalomicin. Antimicrob Agents Chemother 42: 2668-2673.
- Bonnet B, Soullez D, Davioud-Charvet E, Landry
V, Horvath D, Sergheraert C 1997. New spermine and spermidine derivatives
as potent inhibitors of Trypanosoma cruzi trypanothione reductase.
Bioorg Med Chem 5: 1249-1256.
- Bonse S, Richards JM, Ross SA, Lowe G, Krauth-Siegel
RL 2000. (2,2':6',2"-Terpyridine)platinum(II) complexes are irreversible
inhibitors of Trypanosoma cruzi trypanothione reductase but not of
human glutathione reductase. J Med Chem 43: 4812-4821.
- Brener Z 1961. Atividade terapêutica
do 5–nitrofuraldeido- semicarbazona (nitrofurazona) em esquemas de duração
prolongada na infecção experimental pelo Trypanosoma cruzi.
Rev Inst Med Trop São Paulo 3: 43-49.
- Brener Z 1968. Terapêutica experimental
da doença de Chagas. In JR Cançado, Doença de Chagas.
Belo Horizonte, Imprensa Oficial de Minas Gerais, Minas Gerais, p. 510-516.
- Brener Z 1975. Chemotherapy of Trypanosoma
cruzi infection. Adv Pharmacol Chemother 13: 1-44.
- Brener Z 1979. Present status of chemotherapy
and chemoprofilaxis of human trypanosomiasis in the Western hemisphere. PharmacTherap
7: 71-90.
- Brener Z 1984. Recent advances in chemotherapy
of Chagas disease. Mem Inst Oswaldo Cruz 79 (Suppl): 149-155.
- Brener Z 2000. Terapêutica experimental
na doença de Chagas. In Z Brener, Z Andrade, M Barral-Netto (eds),
Trypanosoma cruzi e Doença de Chagas, 2a ed.,
Guanabara Koogan, Rio de Janeiro, p. 379-388.
- Brener Z, Andrade Z, M Barral-Netto 2000.
Trypanosoma cruzi e doença de Chagas, 2a
ed., Guanabara Koogan, Rio de Janeiro, 431 pp.
- Brener Z, Tafuri WL, Maria TA 1969. An electron
microscope study on Trypanosoma cruzi intracellular forms in mice treated
with active nitrofuran compound. Rev Inst Med Trop São Paulo 11:
245-249.
- Brener Z, Cançado JR, Galvão
LM, Da Luz ZM, Filardi LS, Pereira ME, Santos LM, Cançado CB 1993.
An experimental and clinical assay with ketoconazole in the treatment of Chagas
disease. Mem Inst Oswaldo Cruz 88: 149-153.
- Brengio SD, Belmonte SA, Guerreiro E, Giordano
OS, Pietrobon EO, Sosa MA 2000. The sesquiterpene lactone dehydroleucodine
(DhL) affects the growth of cultured epimastigotes of Trypanosoma cruzi.
J Parasitol 86: 407-412.
- Bressi JC, Verlinde CL, Aronov AM, Shaw ML,
Shin SS, Nguyen LN, Suresh S, Buckner FS, Van Voorhis WC, Kuntz ID, Hol WG,
Gelb MH 2001. Adenosine analogues as selective inhibitors of glyceraldehyde-3-phosphate
dehydrogenase of trypanosomatidae via structure-based drug design. J Med
Chem 44: 2080-2093.
- Britto C, Cardoso MA, Vanni CMM, Haslocher-Moreno
A, Xavier SS, Wincker P 1995. Polymerase chain reaction detection of Trypanosoma
cruzi in human blood samples as a tool for diagnosis and treatment evaluation.
Parasitology 110: 241-247.
- Britto C, Silveira C, Cardoso MA, Marques
P, Luquetti A, Macedo V, Fernandes O 2001. Parasite persistence in treated
chagasic patients revealed by xenodiagnosis and polymerase chain reaction.
Mem Inst Oswaldo Cruz 96: 823-826.
- Bronia DH, Pereira BM, Lujan HD, Fretes RE,
Fernandez A, Paglini PA 1999. Ganglioside treatment of acute Trypanosoma
cruzi infection in mice promotes long-term survival and parasitological
cure. Ann Trop Med Parasitol 93: 341-350.
- Bsiri N, Johnson C, Kayirere M, Galy AM, Galy
JP, Barbe J, Osuna A, Mesa-Valle MC, Castilla Calvente JJ, Rodriguez-Cabezas
MN 1996. Trypanocidal structure-activity relationship in 9-thioalkylacridines.
Ann Pharm Fr 54: 27-33.
- Buckner FS, Griffin JH, Wilson AJ, Van Voorhis
WC 2001. Potent anti-Trypanosoma cruzi activities of oxidosqualene
cyclase inhibitors. Antimicrob Agents Chemother 45: 1210-1215.
- Buckner FS, Wilson AJ, White TC, Van Voorhis
WC 1998. Induction of resistance to azole drugs in Trypanosoma cruzi.
Antimicrob Agents Chemother 42: 3245-3250.
- Caceres A, Lopez B, Gonzalez S, Berger I,
Tada I, Maki J 1998. Plants used in Guatemala for the treatment of protozoal
infections. I. Screening of activity to bacteria, fungi and American trypanosomes
of 13 native plants. J Ethnopharmacol 62: 195-202.
- Camargo ME 1966. Fluorescent antibody test
for serodiagnosis of American Trypanosomiasis. Technical modification employing
preserved culture forms of Trypanosoma cruzi in a slide test. Rev
Inst Med Trop São Paulo 8: 227-234.
- Cançado JR 1963. Aspectos clínicos
na padronização dos métodos de avaliação
terapêutica na doença de Chagas. Rev Goiana Med 9 (Supl.):
212-232.
- Cançado JR 1968. Tratamento da doença
de Chagas. In JR Cançado, Doença de Chagas, Imprensa
Oficial de Minas Gerais, Minas Gerais, p. 517-540.
- Cançado JR 1981. Standartization of
protocols for chemotherapy of Chagas disease (Working paper for discussion)
Workshop on Standartizacion of Protocols for the Chemotherapy of Chagas Disease
UNDP/World Bank/TDR. PAHO, Washington, November, 22-26.
- Cançado JR 1985. Tratamento específico.
In JR Cançado, M Chuster (eds), Cardiopatia Chagásica.
Imprensa Oficial de Minas Gerais, Minas Gerais, p. 327-355.
- Cançado JR 1997. Terapêutica
específica. In JCP Dias, JR Coura, Clínica e Terapêutica
da Doença de Chagas. Uma Abordagem Prática para o Clínico
Geral. Fiocruz, Rio de Janeiro, p. 323-351.
- Cançado JR 1999. Criteria of Chagas
disease cure. Mem Inst Oswaldo Cruz 94 (Suppl I): 331-335.
- Cançado JR 2000. Tratamento etiológico
da doença de Chagas. In Z Brener, Z Andrade, M Barral-Netto (eds),
Trypanosoma cruzi e Doença de Chagas, 2a ed.,
Guanabara Koogan, Rio de Janeiro, p. 389-405.
- Cançado JR, Brener Z 1979. Terapêutica.
In Z Brener, Z Andrade (eds), Trypanosoma cruzi e Doença de Chagas,
Guanabara Koogan, Rio de Janeiro, p. 362-424
- Cançado JR, Marra UD, Brener Z 1964.
Ensaio terapêutico clínico com a 5-nitro-2-furaldeido-semicarbazona
(Nitrofurazona) na forma crônica da doença de Chagas. Rev
Inst Med Trop São Paulo 6: 12-16.
- Cançado JR, Marra UD, Lopes M, Mourão
O, Faria CAF, Álvares JM, Salgado AA 1969. Toxicidade y valor terapêutico
del Bay 2502 en la enfermedad de Chagas in tres esquemas posológicos.
Bol Chil Parasitol 24: 28-32.
- Cançado JR, Duval Marra U, Mourão
OG, Álvares JM, Oliveira J, Salgado A 1973. Bases para avaliação
do tratamento específico da doença de Chagas humana segundo
a parasitemia. Rev Soc Bras Med Trop 7: 155-166.
- Cançado JR, Salgado AA, Marra UD, Alvares
JM, Machado JR 1975. Clinical therapeutic trial in chronic Chagas disease
using nifurtimox in 3 schedules of long duration. Rev Inst Med Trop São
Paulo 17: 111-127.
- Cançado JR, Salgado AA, Batista SM,
Chiari C 1976. Segundo ensaio terapêutico com nifurtimox na doença
de Chagas. Rev Goiana Med 22: 203-233.
- Castilla JJ, Mesa-Valle CM, Sanchez-Moreno
M, Arnedo T, Rosales MJ, Mascaro C, Craciunescu D, Osuna A 1996. In vitro
activity and biochemical effectiveness of new organometallic complexes of
osmium(III) against Leishmania donovani and Trypanosoma cruzi. Arznein
Forsch 46: 990-996.
- Cazzulo JJ, Stoka V, Turk V 2001. The major
cysteine proteinase of Trypanosoma cruzi: a valid target for chemotherapy
of Chagas disease. Curr Pharm Des 7: 1143-1156.
- Cerecetto H, Di Maio R, Gonzalez M, Risso
M, Saenz P, Seoane G, Denicola A, Peluffo G, Quijano C, Olea-Azar C 1999.
1,2,5-Oxadiazole N-oxide derivatives and related compounds as potential antitrypanosomal
drugs: structure-activity relationships. J Med Chem 42: 1941-1950.
- Cerecetto H, Di Maio R, Gonzalez M, Risso
M, Sagrera G, Seoane G, Denicola A, Peluffo G, Quijano C, Stoppani AO, Paulino
M, Olea-Azar C, Basombrio MA 2000. Synthesis and antitrypanosomal evaluation
of E-isomers of 5-nitro-2-furaldehyde and 5-nitrothiophene-2-carboxaldehyde
semicarbazone derivatives. Structure-activity relationships. Eur J Med
Chem 35: 343-350.
- Cerisola JA, Rabionvich A, Alvarez M, Corleto
CA, Prumeda J 1972. Enfermedad de Chagas y la transfusión de sangue.
Bol Of Sanit Panam 73: 203-221.
- Cerisola JA, Silva NN, Prata A, Schenone H,
Rohwedder R 1977. Evaluación mediante xenodiagnostico de la efectividad
del nifurtimox en la infección chagásica crónica humana.
Bol Chil Parasitol 32: 51-62.
- Chagas C, Chagas E 1935. Manual de Doenças
Tropicaes e Infectuosas, Vol. I, Editora Freitas Bastos, Rio de Janeiro,
189 pp.
- Chan C, Yin H, Garforth J, McKie JH, Jaouhari
R, Speers P, Douglas KT, Rock PJ, Yardley V, Croft SL, Fairlamb AH 1998. Phenothiazine
inhibitors of trypanothione reductase as potential antitrypanosomal and antileishmanial
drugs. J Med Chem 41: 148-156.
- Chataing B, Concepcion JL, Lobaton R, Usubillaga
A 1998. Inhibition of Trypanosoma cruzi growth in vitro by Solanum
alkaloids: a comparison with ketoconazole. Planta Med 64: 31-36.
- Chiari E, Brener Z 1966. Contribuição
ao diagnóstico parasitológico da doença de Chagas na
fase crônica. Rev Inst Med Trop São Paulo 8: 134-138.
- Chiari E, Dias JCP, Lana M, Chiari CA 1989.
Hemocultures for the parasitological diagnosis of human chronic Chagas disease.
Rev Soc Bras Med Trop 22: 19-23.
- Chiari E, Oliveira AB, Prado MA, Alves RJ,
Galvão LM, Araújo FG 1996. Potential use of WR6026 as prophylaxis
against transfusion-transmitted American trypanosomiasis. Antimicrob Agents
Chemother 40: 613-615.
- Chowdhury SF, Di Lucrezia R, Guerrero RH,
Brun R, Goodman J, Ruiz-Perez LM, Pacanowska DG, Gilbert IH 2001. Novel inhibitors
of leishmanial dihydrofolate reductase. Bioorg Med Chem Lett 11:
977-980.
- Chung MC, Gonçalves MF, Colli W, Ferreira
EI, Miranda MT 1997. Synthesis and in vitro evaluation of potential
antichagasic dipeptide prodrugs of primaquine. J Pharm Sci 86: 1127-1131.
- Cinque GM, Szajnman SH, Zhong L, Docampo R,
Schvartzapel AJ, Rodriguez JB, Gros EG 1998. Structure-activity relationship
of new growth inhibitors of Trypanosoma cruzi. J Med Chem 41: 1540-1554.
- Clark AM 1996. Natural products as a resource
for new drugs. Pharmaceutical Res 13:1133-1141.
- Costa Silva R, Santiago CM, Pontes AL, Andrade
SG 1990. Isoenzymatic pattern of Trypanosoma cruzi Y strain after specific
chemotherapy. Mem Inst Oswaldo Cruz 84: 81-86.
- Coura JR 1996. Perspectivas actuales del trataamiento
específico de la enfermedad de Chagas. Bol Chil Parasitol 51:
69-75.
- Coura JR, Silva JR 1961. Aspectos atuais do
tratamento da doença de Chagas. Rev Bras Med 51: 283-290.
- Coura JR, Ferreira LF, Saad EA, Mortel RE,
Silva JR 1961. Tentativa terapêutica com a nitrofurazona (Furacin) na
forma crônica da doença de Chagas. O Hospital 60: 425-429.
- Coura JR, Ferreira LF, Silva JR 1962. Experiências
com nitrofurazona na fase crônica da doença de Chagas. O Hospital
62: 957-964.
- Coura JR, Brindeiro PJ, Ferreira I 1978. Benznidazole
in the treatment of Chagas disease. Current Chemotherapy. Proc 10 th Int
Cong Chemotherapy 1: 161-162.
- Coura JR, de Abreu LL, Willcox HP, Petana
W 1991. Evaluation of the xenodiagnosis of chronic Chagas patients infected
ten years or over in an area where transmission has been interrupted-Iguatama
and Pains, West Minas Gerais State, Brazil. Mem Inst Oswaldo Cruz 86:
395-398.
- Coura JR, de Abreu LL, Willcox HP, Petana
W 1997. Comparative controlled study on the use of benznidazole, nifurtimox
and placebo, in the chronic form of Chagas disease, in a field area with interrupted
transmission. I. Preliminary evaluation. Rev Soc Bras Med Trop 30:
139-144.
- Croft SL 1994. A rationale for antiparasite
drug discovery. Parasitol Today 10: 385-386.
- Croft SL, Snowdon D, Yardley V 1996. The activities
of four anticancer alkyllysophospholipids against Leishmania donovani,
Trypanosoma cruzi and Trypanosoma brucei. J Antimicrob Chemother 38:
1041-1047.
- Dantas AP 2000. Efeito de agentes anti-microtúbulos
e de derivados de naftoquinonas sobre Trypanosoma cruzi, MSc Thesis, Instituto
Oswaldo Cruz-Fiocruz, Rio de Janeiro, xiv + 204 pp.
- Dantas AP, Ortigão M, Traub-Czebo Y,
Polaquevitch PFC, Barbosa HS, De Castro SL 1998. Studies on the effect of
dinitroaniline herbicides in Trypanosoma cruzi. Mem Inst Oswaldo
Cruz 93 (suppl II): 311
- De Castro SL 1993. The challenge of Chagas
disease chemotherapy: an update of drugs assayed against Trypanosoma cruzi.
Acta Trop 53: 83-98.
- De Castro 2001. Propolis: Biological and pharmacological
activities. Therapeutic uses of this bee-product. Ann Rev Biol Sci,
in press.
- De Castro SL, Higashi KO 1995. Effect
of different formulations of propolis on mice infected with Trypanosoma
cruzi. J Ethnopharmacol 46: 55-59.
- De Castro SL, Pinto MCFR, Pinto AV 1994. Screening
of natural and synthetic drugs against Trypanosoma cruzi: establishing
a structure/activity relationship. Microbios 78: 83-90.
- De Castro SL, Dantas AP, Salomão K,
Prytzyk E, Pereira AS, Aquino Neto FR, Bankova VS 2001. Trypanocidal activity
and chemical composition of Bulgarian samples of propolis. Proc. 4th
COST-B9 Cong Antiprotozoal Chemother, Lisbon, Portugal.
- De Conti R, Rita RM, de Souza EM, Melo PS,
Haun M, De Castro SL, Durán N 1996. In vitro trypanocidal activities
of a novel series of N, N-dimethyl-2-propen-1-amine derivative. Microbios
85: 83-87.
- De Moura KCG, Emery FS, Neves-Pinto C, Pinto
MCFR, Dantas AP, Salomão K, De Castro SL, Pinto AV 2001. Synthesis
and trypanocidal activity of naphthoquinones isolated from Tabebuia
and heterocyclic derivatives: a review from an interdisciplinary study. J
Braz Chem Soc 12: 325-338.
- Deharo E, Loyevsky M, John C, Balanza E, Ruiz
G, Munoz V, Gordeuk VR 2000. Aminothiol multidentate chelators against Chagas
disease. Exp Parasitol 94: 198-200.
- Di Maio R, Cerecetto H, Seoane G, Ochoa C,
Aran VJ, Perez E, Gomez Barrio A, Muelas S 1999. Synthesis and antichagasic
properties of new 1,2,6-thiadiazin-3, 5-dione 1,1-dioxides and related compounds.
Arznein Forsch 49: 759-763.
- Dias JCP 2000. Epidemiologia. In Z Brener,
Z Andrade, M Barral-Netto (eds), Trypanosoma cruzi e Doença de Chagas,
2a ed., Guanabara Koogan, Rio de Janeiro, p. 48-74.
- Diaz de Toranzo EG, Castro JA, Franke de Cazzulo
BM, Cazzulo JJ 1988. Interaction of benznidazole reactive metabolites with
nuclear and kinetoplastic DNA, proteins and lipids from Trypanosoma cruzi.
Experientia 44: 880-881.
- DoCampo R 2001. Recent developments in the
chemotherapy of Chagas disease. Curr Pharm Design 7: 1157-1164.
- DoCampo R, Moreno SNJ 1986. Free radical metabolism
of antiparasitic agents. Fed Proceed 45: 2471-2476.
- DoCampo R, Moreno SNJ 2001. Bisphosphonates
as chemotherapeutic agents against trypanosomatids and Apicomplexan parasites.
Current Drug Targets-Infectious Disorders 1: 51-61.
- Dubin M, Fernandez Villamil SH, Stoppani AO
2001. Cytotoxicity of beta-lapachone, a naphthoquinone with possible therapeutic
use. Medicina (B Aires) 61: 343-350.
- Eakin AE, Guerra A, Focia PJ, Torres-Martinez
J, Craig SP 1997. Hypoxanthine phosphoribosyltransferase from Trypanosoma
cruzi as a target for structure-based inhibitor design: crystallization
and inhibition studies with purine analogs. Antimicrob Agents Chemother
41: 1686-1692.
- Engel JC, Doyle PS, Palmer J, Bainton DF,
McKerrow JH 1998a. Cysteine protease inhibitors after Golgi complex ultrastructure
and function in Trypanosoma cruzi. J Cell Sci 111: 597-606.
- Engel JC, Doyle PS, Hsieh I, McKerrow JH 1998b.
Cysteine protease inhibitors cure an experimental Trypanosoma cruzi
infection. J Exp Med 188: 725-734.
- Fairlamb AH 1999. Future prospects for the
chemotherapy of Chagas disease. Medicina (B Aires) 59: 179-187.
- Fairlamb AH, Cerami A 1992. Metabolism and
functions of trypanothione in the kinetoplastida. Annu Rev Micorbiol 46:
695-729.
- Fernandez-Gomez R, Zerrouk H, Sebti F, Loyens
M, Benslimane A, Ouaissi MA 1994. Growth inhibition of Trypanosoma cruzi
and Leishmania donovani infantum by different snake venoms: preliminary
identification of proteins from Cerastes cerastes venom which interact
with the parasites. Toxicon 32: 875-882.
- Ferreira HO 1961. Forma aguda da doença
de Chagas tratada pela nitrofurazona. Rev Inst Med Trop São Paulo
3: 287-289.
- Ferreira HO 1962. Fase aguda da doença
de Chagas. O Hospital 61: 307-311.
- Ferreira HO 1976. Ensaio terapêutico-clínico
com benznidazol na doença de Chagas. Rev Inst Med Trop São
Paulo 18: 357-364.
- Ferreira HO 1990. Tratamento da forma indeterminada
da doença de Chagas com nifurtimox e benznidazol. Rev Soc Bras Med
Trop 23: 209-211.
-
Ferreira MS, Nishioka SA, Rocha
A, Silva AM 1997a. Doença de Chagas e imunossupressão. In
JCP Dias, JR Coura (eds), Clínica e Terapêutica da Doença
de Chagas. Fiocruz, Rio de Janeiro, p. 365-379.
-
Ferreira MS, Nishioka SD, Silvestre
MT, Borges AS, Nunes-Araujo FR, Rocha A 1997b. Reactivation of Chagas disease
in patients with AIDS: report of three new cases and review of the literature.
Clin Infect Dis 25: 1397-1400.
-
Fichera L, Esteva M, Wimmer Z,
Rodriguez JB, Gros EG 1995. Effects of juvenile hormone analogues (JHA)
on the development of Trypanosoma cruzi. Z Naturforsch 50C: 578-580.
-
Fournet A, Inchausti A, Yaluff
G, Rojas De Arias A, Guinaudeau H, Bruneton J, Breidenbach MA, Karplus PA,
Faerman CH 1998. Trypanocidal bisbenzylisoquinoline alkaloids are inhibitors
of trypanothione reductase. J Enzym Inhib 13: 1-9.
-
Fournet A, Rojas De Arias A,
Ferreira ME, Nakayama H, Torres de Ortiz S, Schinini A, Samudio M, Vera
de Bilbao N, Lavault M, Bonte F 2000. Efficacy of the biisbenziylisoquinoline
alkaloids in acute and chronic Trypanosoma cruzi murine model. Int
J Antimicrob Agents 13: 189-195.
-
Fragata Filho AA, Boianain E,
Silva MAD, Correia EB, Borges Filho R, Martins C, Salene V, Batlouni M,
Souza E 1995. Validade do tratamento etiológico da fase crônica
da doença de Chagas com benznidazol. Arq Bras Cardiol 65 (Supl
I): 71.
-
Fragoso SP, Mattei D, Hines JC,
Ray D, Goldenberg S 1998. Expression and cellular localization of Trypanosoma
cruzi type II DNA topoisomerase. Mol Biochem Parasitol 94: 197-204.
-
Freymann DM, Wenck MA, Engel
JC, Feng J, Focia PJ, Eakin AE, Craig SP 2000. Efficient identification
of inhibitors targeting the closed active site conformation of the HPRT
from Trypanosoma cruzi. Chem Biol 7: 957-968.
-
Galhardo MCG, Martins, I, Hasslocher-Moreno
A, Xavier SS, Coelho JMC, Vasconcellos ACV, Santos RR 1999. Reactivação
da infecção por Trypanosoma cruzi em paciente com síndrome
de imunodeficiência adquirida. Rev Soc Bras Med Trop 32: 291-294.
-
Girault S, Davioud-Charvet E,
Salmon L, Berecibar A, Debreu MA, Sergheraert C 1998. Structure-activity
relationships in 2-aminodiphenylsulfides against trypanothione reductase
from Trypanosoma cruzi. Bioorg Med Chem Lett 8: 1175-1180.
-
Girault S, Grellier P, Berecibar
A, Maes L, Mouray E, Lemiere P, Debreu MA, Davioud-Charvet E, Sergheraert
C 2000. Antimalarial, antitrypanosomal, and antileishmanial activities and
cytotoxicity of bis(9-amino-6-chloro-2-methoxyacridines): influence
of the linker. J Med Chem 43: 2646-2654.
-
Gonzales-Perdomo M, De Castro
SL, Meirelles MN, Goldenberg S 1990. Trypanosoma cruzi proliferation
and differentiation are blocked by topoisomerase II inhibitors. Antimicrob
Agents Chemother 34: 1707-1714.
-
Gonzalez-Martin G, Figueroa C,
Merino I, Osuna A 2000. Allopurinol encapsulated in polycyanoacrylate nanoparticles
as potential lysosomatropic carrier: preparation and trypanocidal activity.
Eur J Pharm Biopharm 49: 137-142.
-
Gonzalez-Martin G, Merino I,
Rodriguez-Cabezas MN, Torres M, Nunez R, Osuna A, 1998. Characterization
and trypanocidal activity of nifurtimox-containing and empty nanoparticles
of polyethylcyanoacrylates. J Pharm Pharmacol 50: 29-35.
-
Grellier P, Sinou V, Garreau-de
Loubresse N, Bylen E, Boulard Y, Schrevel J 1999. Selective and reversible
effects of Vinca alkaloids on Trypanosoma cruzi epimastigote
forms: blockage of cytokinesis without inhibition of the organelle duplication.
Cell Motil Cytoskeleton 42: 36-47.
-
Henderson GB, Ulrich P, Fairlamb
AH, Rosemberg I, Pereira M, Sela M, Cerami A 1988. "Subversive"
substrates for the enzyme trypanothione disulphide reductase, alternative
approach to chemoterapy of Chagas disease. Proc Natl Acad Sci USA
85: 5374-5378.
-
Higuchi ML, Brito T, Reis M,
Bellotti G, Pereira-Barreto AC, Pileggi F 1993. Correlation between Trypanosoma
cruzi parasitism and myocardial inflammation in human chronic chagasic
myocardits. Light microscopy and immunohistochemical findings. Cardiovasc
Pathol 2: 101-106.
-
Higuchi ML, Reis M, Aiello VD,
Benvenutti LA, Gutierrez PS, Bellotti G, Pileggi F 1997. Human chronic chagasic
myocarditis is Trypanosoma cruzi antigen and CD8+ T cell
dependent. Am J Trop Med Hyg 56: 485-489.
-
Ianni BM, Arteaga E, Mady C,
Barretto ACP, Pileggi F 1993. Uso de benznidazol em chagásicos na
forma indeterminada: Resultados a longo prazo. Arq Bras Cardiol 61 (Supl
II): 130.
-
Jatene AD, Costa R, Jatene MB
1997. Tratamento cirúrgico da cardiopatia chagásica. In JCP
Dias, JR Coura (eds), Clínica e Terapêutica da Doença
de Chagas. Fiocruz, Rio de Janeiro, p. 255-265.
-
Jones EM, Colley DG, Tostes S,
Lopes ER, Venencak-Jones C, Mc Curley TL 1993. Amplification of Trypanosoma
cruzi DNA sequence from inflammatory lesions in human chagasic cardiomyopathy.
Am J Trop Med Hyg 48: 348-357.
-
Junqueira ACV, Chiari E, Wincker
P 1996. Comparison of the polymerase chain reaction with two classical parasitological
methods, for diagnosis of Chagas disease in endemic region of North-Eastern
Brazil. Trans R Soc Trop Med Hyg 90: 129-132.
-
Kerschmann RL, Wolfson JS, McHugh
GL, Dickersin GR, Hooper DC, Swartz MN 1989. Novobiocin-induced ultrastructural
changes and antagonism of DNA synthesis in Trypanosoma cruzi amastigotes
growing in cell-free medium. J Protozool 36: 14-20.
-
Kinnamon KE, Poon BT, Hanson
WL, Waits VB 1996. Primaquine analogues that are potent anti-Trypanosoma
cruzi agents in a mouse model. Ann Trop Med Parasitol 90: 467-474.
-
Kinnamon KE, Poon BT, Hanson
WL, Waits VB 1997. Evidence that certain 8-aminoquinolines are potentially
effective drugs against Chagas disease. Ann Trop Med Parasitol 91:
147-52.
-
Krauth-Siegel RL, Enders B, Henderson
GB, Fairlamb AH, Schirmer RH 1987. Trypanothione reductase from Trypanosoma
cruzi. Purification and characterization of the crystalline enzyme.
Eur J Biochem 164: 123-128.
-
Lalmanach G, Mayer R, Serveau
C, Scharfstein J, Gauthier F 1996. Biotin-labelled peptidyl diazomethane
inhibitors derived from the substrate-like sequence of cystatin: targeting
of the active site of cruzipain, the major cysteine proteinase of Trypanosoma
cruzi. Biochem J 318: 395-399.
-
Lane JE, Ribeiro-Rodrigues R,
Suarez CC, Bogitsh BJ, Jones MM, Singh PK, Carter CE 1996. In vitro
trypanocidal activity of tetraethylthiuram disulfide and sodium diethylamine-N-carbodithioate
on Trypanosoma cruzi. Am J Trop Med Hyg 55: 263-266.
-
Lane JE, Bogitsh BJ, Ribeiro-Rodrigues
R, Kral MV, Jones MM, Carter CE 1998. Ultrastructural effects of the chelating
agent 1,10-phenanthroline on Trypanosoma cruzi epimastigotes in
vitro. Parasitol Res 84: 399-402.
-
Lauria-Pires L, Castro CN, Emanuel
A, Prata A 1988. Ineficácia do allopurinol em pacientes na fase aguda
da doença de Chagas. Rev Soc Med Trop 21: 79
-
Li Z, Fennie MW, Ganem B, Hancock
MT, Kobaslija M, Rattendi D, Bacchi CJ, O'Sullivan MC 2001. Polyamines with
N-(3-phenylpropyl) substituents are effective competitive inhibitors of
trypanothione reductase and trypanocidal agents. Bioorg Med Chem Lett
11: 251-254.
-
Lira R, Contreras LM, Rita RM,
Urbina JA 2001. Mechanism of action of anti-proliferative lysophospholipid
analogues against the protozoan parasite Trypanosoma cruzi: potentiation
of in vitro activity by the sterol biosynthesis inhibitor ketoconazole.
J Antimicrob Chemother 47: 537-546.
-
Lopes JN, Cruz FS, DoCampo R,
Vasconcellos ME, Sampaio MCR, Pinto AV, Gilbert B 1978. In vitro
and in vivo evaluation of the toxicity of 1,4-naphthoquinone
and 1,2-naphthoquinone derivatives against Trypanosoma cruzi. Ann
Trop Med Parasitol 72: 523-531.
-
Lujan HD, Paglini P, Fretes R,
Fernandez A, Fidelio GD, Bronia DH 1993. Effect of gangliosides on Trypanosoma
cruzi infection in mice. Life Sci 53: PL69-73.
-
Luque F, Fernandez-Ramos C, Entrala
E, Rosales MJ, Navarro JA, Romero MA, Salas JM, Sanchez-Moreno M 2000. In
vitro evaluation of newly synthesized [1,2,4]triazolo[1, 5a]pyrimidine
derivatives against Trypanosoma cruzi, Leishmania donovani and Phytomonas
staheli. Comp Biochem Physiol C Toxicol Pharmacol 126:
39-44.
-
Luz ZMP, Coutinho MG, Cançado
JR, Krettli AU 1994. Hemocultura: técnica sensível na detecção
do Trypanosoma cruzi em pacientes chagásicos crônicos.
Rev Soc Bras Med Trop 27: 143-148.
-
Macêdo VO 1997. Forma indeterminada
da doença de Chagas. In JCP Dias, JR Coura (eds), Clínica
e Terapêutica da Doença de Chagas. Uma Abordagem Prática
para o Clínico Geral. Fiocruz, Rio de Janeiro, p. 383-409.
-
Mafezoli J, Vieira PC, Fernandes
JB, Da Silva MF, De Albuquerque S 2000. In vitro activity of Rutaceae
species against the trypomastigote form of Trypanosoma cruzi. J
Ethnopharmacol 73: 335-340.
-
Mahiou V, Roblot F, Fournet A,
Hocquemiller R 2000. Bisbenzylisoquinoline alkaloids from Guatteria boliviana
(Annonaceae). Phytochemistry 54: 709-716.
-
Marcucci MC, Ferreres F, García-Viguera
C, Bankova VS, De Castro SL, Dantas AP, Valente PHM, Paulino N 2001. Phenolic
compounds from Brazilian propolis with pharmacological activities. J
Ethnopharmacol 74: 105-112.
-
Martin MB, Grimley JS, Lewis
JC, Heath HT, 3rd, Bailey BN, Kendrick H, Yardley V, Caldera A, Lira R,
Urbina JA, Moreno SN, Docampo R, Croft SL, Oldfield E 2001. Bisphosphonates
inhibit the growth of Trypanosoma brucei, Trypanosoma cruzi, Leishmania
donovani, Toxoplasma gondii, and Plasmodium falciparum: A potential
route to chemotherapy. J Med Chem 44: 909-916.
-
Maya JD, Morello A, Repetto Y,
Tellez R, Rodriguez A, Zelada U, Puebla P, Caballero E, Medarde M, Nunez-Vergara
LJ, Squella JA, Bonta M, Bollo S, San Feliciano A 2000. Effects of 3-chloro-phenyl-1,4-dihydropyridine
derivatives on Trypanosoma cruzi epimastigotes. Comp Biochem Physiol
C Pharmacol Toxicol Endocrinol 125: 103-109.
-
Mazza S, Cossio R, Zucardi E
1937. Primer caso agudo de enfermedad de Chagas, comprovado em Tucuman y
su tratamiento com Bayer 7602. Mis Estudios Patolog Reg Argentina Publ
70 (Univ Buenos Aires) (MEPRA) 32: 3-18.
-
Mazza S, Basso G, Basso R 1942.
Ensayos terapéuticos del produto 9736 (As) Bayer y de su acción
comparada en el 7602 (Ac) Bayer en la enfermedad de Chagas. Mis Estudios
Patolog Reg Argentina Publ 70 (Univ Buenos Aires) (MEPRA) 61: 1-76.
-
Medrano NM, Luz MRMP, Cabello
P, Tapia GT, Van Leuven F, Araújo-Jorge TC 1996. Acute Chagas disease:
Plasma levels of alpha-2-macroglobulin and C-reactive protein in children
under 13 years in a high endemic area of Bolivia. J Trop Pediatr 42:
68-74.
-
Molina J, Martins-Filho O, Brener
Z, Romanha AJ, Loebenberg D, Urbina JA 2000. Activities of the triazole
derivative SCH 56592 (posaconazole) against drug-resistant strains of the
protozoan parasite Trypanosoma (Schizotrypanum) cruzi in immunocompetent
and immunosuppressed murine hosts. Antimicrob Agents Chemother 44:
150-155.
-
Molina J, Urbina J, Gref R, Brener
Z, Rodrigues Junior JM 2001. Cure of experimental Chagas disease by the
bis-triazole DO870 incorporated into "stealth" polyethyleneglycol-polylactide
nanospheres. J Antimicrob Chemother 47: 101-104.
-
Montalvetti A, Bailey BN, Martin
MB, Severin GW, Oldfield E, DoCampo R 2001. Bisphosphonates are potent inhibitors
of Trypanosoma cruzi farnesyl pyrophosphate synthase. J Biol Chem
276: 33930-33937.
-
Morello A, Lipchenca I, Cassels
BK, Speisky H, Aldunate J, Repetto Y 1994. Trypanocidal effect of boldine
and related alkaloids upon several strains of Trypanosoma cruzi. Comp
Biochem Physiol Pharmacol Toxicol Endocrinol 107: 367-371.
-
Morello A, Pavani M, Garbarino
JA, Chamy MC, Frey C, Mancilla J, Guerrero A, Repetto Y, Ferreira J 1995.
Effects and mode of action of 1,4-naphthoquinones isolated from Calceolaria
sessilis on tumoral cells and Trypanosoma parasites. Comp
Biochem Physiol C Pharmacol Toxicol Endocrinol 112: 119-128.
-
Muelas S, Di Maio R, Cerecetto
H, Seoane G, Ochoa C, Escario JA, Gomez-Barrio A 2001. New thiadiazine derivatives
with activity against Trypanosoma cruzi amastigotes. Folia Parasitol
(Praha) 48: 105-108.
-
Muelas-Serrano S, Nogal JJ, Martinez-Diaz
RA, Escario JA, Martinez-Fernandez AR, Gomez-Barrio A 2000. In vitro
screening of American plant extracts on Trypanosoma cruzi and Trichomonas
vaginalis. J Ethnopharmacol 71: 101-107.
-
Neira I, Poblete L, Porcille
P, Silva P, Araya J, Borquez J, Morales G, Loyola LA, Sagua H 1998. Activity
of Dipernoids isolated from Azorella compacta (Llareta) on Trypanosoma
cruzi amastigotes. Bol Chil Parasitol 53: 9-13.
-
Neves-Pinto C, Dantas AP, De
Moura KCG, Emery FS, Polequevitch PF, Pinto MCFR, De Castro SL, Pinto AV
2000. Chemical reactivity studies with naphthoquinones from Tabebuia
with anti-trypanosomal efficacy. Arzneim Forsch 50: 1120-1128.
-
Nunez-Vergara LJ, Squella JA,
Bollo-Dragnic S, Morello A, Repetto Y, Aldunate J, Letelier ME 1997. Nitro
aryl 1,4-dihydropyridine derivatives: effects on Trypanosoma cruzi. Comp
Biochem Physiol C Pharmacol Toxicol Endocrinol 118: 105-111.
-
Nussenzweig V, Sonntag R, Biancalana
A, Freitas JLP, Amato Neto V, Kloetzel J 1953. Ação da violeta
de genciana sobre o T. cruzi in vitro: sua importância na esterilização
do sangue destinado à transfusão. Rev Paul Med 42:
57-58.
-
Ochoa C, Perez E, Perez R, Suarez
M, Ochoa E, Rodriguez H, Gomez Barrio A, Muelas S, Nogal JJ, Martinez RA
1999. Synthesis and antiprotozoan properties of new 3,5-disubstituted-tetrahydro-2H-1,3,5-thiadiazine-2-thione
derivatives. Arznein Forsch 49: 764-769.
-
Oliveira DA, Fernandes AM, De
Conti R, Rodriguez JA, Haun M, Souza-Brito AR, De Castro SL, Durán
N 1999. Evaluation of in vitro toxicity of N,N-dimethyl-2-propen-1-amines
isomers. Pharmazie 54: 847-850.
-
Olmo E, Armas MG, Lopez-Perez
JL, Ruiz G, Vargas F, Gimenez A, Deharo E, Feliciano AS 2001. Anti-Trypanosoma
activity of some natural stilbenoids and synthetic related heterocyclic
compounds. Bioorg Med Chem Lett 11: 755-757.
-
Paglini-Oliva P, Fernandez AR,
Fretes R, Peslman A 1998. Structural, ultrastructural studies and evolution
of Trypanosoma cruzi-infected mice treated with thioridazine. Exp
Mol Pathol 65: 78-86.
-
Pereira DG, De Castro SL, Durán
N 1998. Activity of N,N-dimethyl-1-2-propen-1-amine derivatives in mice
experimentally infected with Trypanosoma cruzi. Acta Trop 69: 205-211.
-
Pinto AV, Neves Pinto C, Pinto
MCFR, Santa-Rita RM, Pezzella C, De Castro SL 1997. Trypanocidal activity
of synthetic heterocyclic derivatives from active quinones from Tabebuia
sp. Arzneim-Forsch 4: 74-79.
-
Prata A, Macedo V, Santos I,
Cerisola JA, Silva N 1975. Tratamento da doença de Chagas pelo nifurtimox
(Bayer 2505). Rev Soc Brasil Med Trop 6: 297-308.
-
Rassi A, Luquetti AO 1992. Therapy
of Chagas disease. In S Wendel, Z Brener, E Camargo, A Rassi (eds), Chagas
Disease (American Trypanosomiasis): its Impact on Transfusion and Clinical
Medicine. ISBT, São Paulo, p. 237-247.
-
Rassi A, Amato Neto V, De Siqueira
AF, Ferriolli Filho F, Amato VS, Rassi Jr A 1999. Protective effect of benznidazole
against parasite reactivation in patients chronically infected with Trypanosoma
cruzi and treated with corticoids for associated diseases. Rev Soc
Bras Med Trop 32: 475-482.
-
Reche P, Arrebola R, Santi DV,
Gonzalez-Pacanowska D, Ruiz-Perez LM 1996. Expression and characterization
of the Trypanosoma cruzi dihydrofolate reductase domain. Mol Biochem
Parasitol 76: 175-185.
-
Rivarola HW, Fernandez AR, Enders
JE, Fretes R, Gea S, Suligoy M, Palma JA, Paglini-Oliva P 1999. Thioridazine
treatment modifies the evolution of Trypanosoma cruzi infection in
mice. Ann Trop Med Parasitol 93: 695-702.
-
Rivas P, Cassels BK, Morello
A, Repetto Y 1999. Effects of some b-carboline
alkaloids on intact Trypanosoma cruzi epimastigotes. Comp Biochem
Physiol C Pharmacol Toxicol Endocrinol 122: 27-31.
-
Rodrigues RR, Lane JE, Carter
CE, Bogitsh BJ, Singh PK, Zimmerman LJ, Molenda JJ, Jones MM 1995. Chelating
agent inhibition of Trypanosoma cruzi epimastigotes in vitro.
J Inorg Biochem 60: 277-288.
-
Rodriguez JB, DoCampo R, Gros
EG 2000. Sulphur-containing derivatives structurally related to fenoxycarb
are potent growth inhibitors against the intracellular form of Trypanosoma
cruzi. Int J Antimicrob Agents 13: 215-218.
-
Roush WR, Hernandez AA, McKerrow
JH, Selzer PM, Hansell E, Engel JC 2000. Design, synthesis and evaluation
of D-homophenylalanylepoxysuccinate inhibitors of the trypanosomal cystein
protease cruzain. Tetrahedron 56: 9747-9762.
-
Rovai LE, Aoki A, Gerez de Burgos
NM, Blanco A 1990. Effect of gossypol on trypomastigotes and amastigotes
of Trypanosoma cruzi. J Protozool 37: 280-286.
-
Ryley JF, McGregor S, Wilson
RG 1988. Activity of ICI 195,739 - a novel orally active bistriazole - in
rodent models of fungal and protozoal infection. Ann N Y Acad Sci
310: 328.
-
Salmon-Chemin L, Buisine E, Yardley
V, Kohler S, Debreu MA, Landry V, Sergheraert C, Croft SL, Krauth-Siegel
RL, Davioud-Charvet E 2001. 2- and 3-substituted 1,4-naphthoquinone derivatives
as subversive substrates of trypanothione reductase and lipoamide dehydrogenase
from Trypanosoma cruzi: synthesis and correlation between redox cycling
activities and in vitro cytotoxicity. J Med Chem 44: 548-565.
-
Santa-Rita RM, Barbosa HS, Meirelles
MNL, De Castro SL 2000. Effect of alkyllysophospholipids on the proliferation
and differentiation of Trypanosoma cruzi. Acta Trop 75:
219-228.
-
Santos AF, Ferraz PA, de Abreu
FC, Chiari E, Goulart MO, Sant'Ana AE 2001. Molluscicidal and trypanocidal
activities of lapachol derivatives. Planta Med 67: 92-3.
-
Schenone H, Concha L, Aranda
R, Rojas A, Alfaro E 1969. Experiência terapêutica con el Bayer
2502 en la infección chagasica cronica del adulto. Importancia del
uso adequado del xenodiagnóstico. Bol Chil Parasitol 24: 66-69.
-
Schenone H, Concha L, Aranda
R, Rojas A, Alfaro E, Knierin E, Rojo M 1975. Atividade quimioterápica
de um derivado nitroimidazolacetamida na infecção chagásica
crônica. Bol Chil Parasitol 30: 91-93.
-
Schenone H, Concha L, Aranda
R, Rojas A, Knierim F, Rojo M 1972. Treatment of chronic Chagas infection
with Lampit. Bol Chil Parasitol 27: 11-14.
-
Schenone H, Rojas A, Alfaro E,
Concha L, Aranda R 1981. Estudio longitudinal de la persistencia de la acción
terapéutica del nifurtimox y del benznidazol en pacientes con infección
chagásica crónica. Bol Chil Parasitol 36: 59-62.
-
Schmeda-Hirschmann G, Astudillo
L, Bastida J, Codina C, Rojas de Arias A, Ferreira ME, Inchaustti A, Yaluff
G 2001. Cryptofolione derivatives from Cryptocarya alba fruits. J
Pharm Pharmacol 53: 563-567.
-
Schmunis GA 2000. A tripanosomíase
americana e seu impacto na saúde pública das américas.
In Z Brener, Z Andrade, M Barral-Netto (eds), Trypanosoma cruzi e Doença
de Chagas, 2a ed., Guanabara Koogan, Rio de Janeiro,
p. 1-15.
-
Silva NN, Kuhn G, Santos JFC,
Von Eye G, Chaer JAB 1974. Eficácia e tolerância do nitrofurfurilidene
na fase crônica da moléstia de Chagas. Rev Soc Bras Med
Trop 88: 325-334.
-
Silveira JF 1992. Trypanosoma
cruzi recombinant antigens for serodiagnosis. In S Wendel, Z Brener,
E Camargo, A Rassi (eds), Chagas Disease (American Trypanosomiasis):
its Impact on Transfusion and Clinical Medicine. ISBT, São Paulo,
p. 207-218.
-
Singh PK, Jones MM, Lane JE,
Nesset A, Zimmerman LJ, Ribeiro-Rodrigues R, Richter A, Stenger MR, Carter
CE 1997. Synthesis and in vitro trypanocidal activity of some novel iron
chelating agents. Arznein Forsch 47: 311-315.
-
Solari A, Saavedra H, Sepulveda
C, Oddó D, Acuña G, Labarca J, Muñoz S, Cuny G, Brengues
C, Veas F, Bryan RT 1993. Successful treatment of Trypanosoma cruzi
encephalitis in a patient with hemophilia and AIDS. Clin Inf Dis 16:
255-259.
-
Sosa Estani S, Segura EL, Ruiz
AM, Velazquez E, Porcel BM, Yampotis C 1998. Efficacy of chemotherapy with
benznidazole in children in the indeterminate phase of Chagas disease. Am
J Trop Med Hyg 59: 526-529.
-
Souza DH, Garratt RC, Araújo
AP, Guimarães BG, Jesus WD, Michels PA, Hannaert V, Oliva G 1998.
Trypanosoma cruzi glycosomal glyceraldehyde-3-phosphate dehydrogenase:
structure, catalytic mechanism and targeted inhibitor design. FEBS Lett
424: 131-135.
-
Stoka V, Nycander M, Lenarcic
B, Labriola C, Cazzulo JJ, Bjork I, Turk V 1995. Inhibition of cruzipain,
the major cysteine proteinase of the protozoan parasite, Trypanosoma
cruzi, by proteinase inhibitors of the cystatin superfamily. FEBS
Lett 370: 101-104.
-
Storino R, Galleano R, Sosa R
1994. Tratamento antiparasitário específico. In R Storino,
J Milei (eds), Enfermedad de Chagas, Mosby Doyma, Argentina, p. 557-568.
-
Sturm NR, Degrave W, Morel CM,
Simpson L 1989. Sensitive detection by amplification of kinetoplast minicircle
DNA sequences: use in diagnosis of Chagas disease. Mol Biochem Parasitol
33: 205-214.
-
Sundar S, Goyal AK, More DK,
Singh MK, Murray HW 1998. Treatment of antimony-unresponsive Indian visceral
leishmaniasis with ultra-short courses of amphotericin-B-lipid complex.
Ann Trop Med Parasitol 92: 755-764.
-
Sundar S, Makharia A, More DK,
Agrawal G, Voss A, Fischer C, Bachmann P, Murray HW 2000. Short-course of
oral miltefosine for treatment of visceral leishmaniasis. Clin Infect
Dis 31: 1110-1113.
-
Szajnman SH, Yan W, Bailey BN,
DoCampo R, Elhalem E, Rodriguez JB 2000. Design and synthesis of aryloxyethyl
thiocyanate derivatives as potent inhibitors of Trypanosoma cruzi
proliferation. J Med Chem 43: 1826-1840.
-
Teixeira AR, Cordoba JC, Souto
Maior IC, Solorzano E 1990. Chagas disease: lymphoma growth in rabbits treated
with benznidazole. Am J Trop Med Hyg 43: 146-158.
-
Tomazela DM, Pupo MT, Passador
EA, da Silva MF, Vieira PC, Fernandes JB, Fo ER, Oliva G, Pirani JR 2000.
Pyrano chalcones and a flavone from Neoraputia magnifica and their
Trypanosoma cruzi glycosomal glyceraldehyde-3-phosphate dehydrogenase-inhibitory
activities. Phytochemistry 55: 643-651.
-
Tomimori-Yamashita J, Deps PD,
Almeida DR, Enokihara MM, De Seixas MT, Freymuller E 1997. Cutaneous manifestation
of Chagas disease after heart transplantation: successful treatment with
allopurinol. Br J Dermatol 37: 626-630.
-
Traub-Cseko YM, Ramalho-Ortigao
JM, Dantas AP, De Castro SL, Barbosa HS, Downing KH 2001. Dinitroaniline
herbicides against protozoan parasites: the case of Trypanosoma cruzi.
Trends Parasitol 17: 136-141.
-
Umezawa ES, Nascimento MS, Kesper
Jr, Coura JR, Borges-Pereira J, Junqueira ACV, Camargo ME 1996. Immunoblot
assay using excreted-secreted antigens of Trypanosoma cruzi in serodiagnosis
of congenital, acute and chronic Chagas disease. J Clin Microbiol 37:
1554-1560.
-
Umezawa ES, Nascimento MS, Stolf
AMS 2001. Enzime-linked immunosorbent assay with Trypanosoma cruzi
excreted-secreted antigens (TESA-ELISA) for serodiagnosis of acute and chronic
Chagas' disease. Diag Microbiol Infect Disease 39: 169-176.
-
Urbina JA 1999. Chemotherapy
of Chagas disease: the how and the why. J Mol Med 77: 332-338.
-
Urbina JA, Payares G, Molina
J, Sanoja C, Liendo A, Lazardi K, Piras MM, Piras R, Perez N, Wincker P,
Ryley JF 1996. Cure of short- and long-term experimental Chagas disease
using D0870. Science 273: 969-971.
-
Urbina JA, Moreno B, Vierkotter
S, Oldfield E, Payares G, Sanoja C, Bailey BN, Yan W, Scott DA, Moreno SN,
DoCampo R 1999. Trypanosoma cruzi contains major pyrophosphate stores,
and its growth in vitro and in vivo is blocked by pyrophosphate
analogs. J Biol Chem 274: 33609-33615.
-
Urbina JA, Lira R, Visbal G,
Bartroli J 2000. In vitro antiproliferative effects and mechanism
of action of the new triazole derivative UR-9825 against the protozoan parasite
Trypanosoma (Schizotrypanum) cruzi. Antimicrob Agents Chemother 44:
2498-2502.
-
Viotti R, Vigliano C, Armenti
H, Segura E 1994. Treatment of chronic Chagas disease with benznidazole:
clinical and serologic evolution of patients with long-term follow-up. Am
Heart J 127: 151-162.
-
Voller A, Draper C, Bidwell DE,
Bartlett A 1975. A micro-plate enzyme-linked immunosorbent assay (ELISA)
for Chagas disease. The Lancet 1: 426-429.
-
Waechter AI, Yaluff G, Inchausti
A, Rojas de Arias A, Hocquemiller R, Cavé A, Fournet A 1998. Leishmanicidal
and trypanocidal activities of acetogenins isolated from Annona glauca.
Phytotherapy Res 12: 541-544.
-
Wendel S, Dias JCP 1992. Transfusion
transmitted Chagas disease. In S Wendel, Z Brener, E Camargo, A Rassi (eds),
Chagas Disease (American trypanosomiasis): its Impact on Transfusion
and Clinical Medicine. ISBT, São Paulo, p. 103-134.
-
Wieder T, Reutter W, Orfanos
CE, Geilen CC 1999. Mechanisms of action of phospholipid analogs as anticancer
compounds. Prog Lipid Res 38: 249-259.
-
Winker P, Britto C, Borges Pereira,
Cardoso MA, Coleman W, Morel CM 1994. Use of simplified polymerase chain
reaction procedures to detect, Trypanosoma cruzi in blood samples
from chronic chagasic patients in rural area. Am J Trop Med Hyg 51:
771-777.
-
Yong V, Schmitz V, Vannier-Santo
MA, de Lima AP, Lalmanach G, Juliano L, Gauthier F, Scharfstein J 2000.
Altered expression of cruzipain and a cathepsin B-like target in a Trypanosoma
cruzi cell line displaying resistance to synthetic inhibitors of cysteine-proteinases.
Mol Biochem Parasitol 109: 47-59.
-
Zaidenberg A, Tournier H, Schinella
G, Marín G, Buschiazzo H 1999. Effects of trifluralin on Trypanosoma
cruzi in vitro and in vivo. Pharmacol Toxicol 84:
98-100.
-
Zuccotto F, Brun R, Gonzalez
Pacanowska D, Ruiz Perez LM, Gilbert IH 1999. The structure-based design
and synthesis of selective inhibitors of Trypanosoma cruzi dihydrofolate
reductase. Bioorg Med Chem Lett 9: 1463-1468.
- Zuccotto F, Martin AC, Laskowski RA, Thornton
JM, Gilbert IH 1998. Dihydrofolate reductase: a potential drug target in
trypanosomes and leishmania. J Comput Aided Mol Des 12: 241-257.
© 2002
Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02002f1.jpg]
|

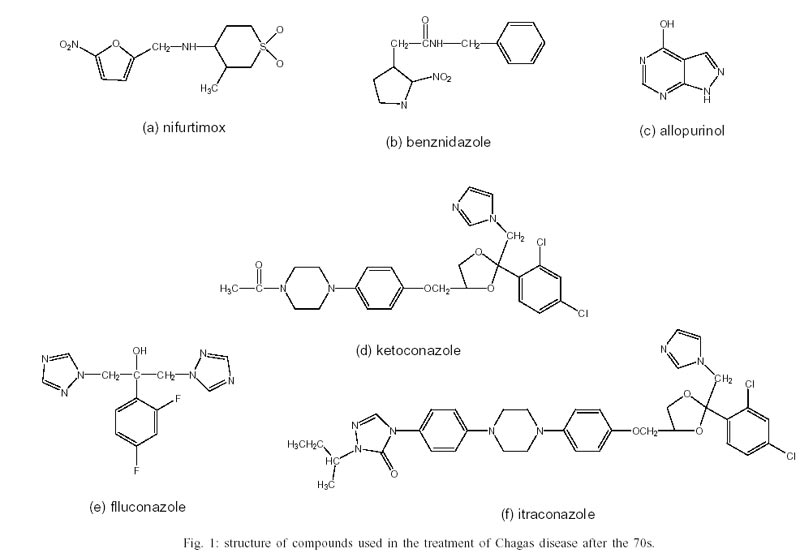{kind=link}