
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 1, 2002, pp. 65-72
|
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(1) 2002, pp. 65-72
A Cladistic Analysis of Inaequalium
(Coscarón & Wygodzinsky, 1984), with Information on Geographical Distribution
(Diptera: Simuliidae)
MN Strieder/+, V Py-Daniel*
Laboratório de Entomologia, Centro de Ciências
da Saúde, Universidade do Vale do Rio dos Sinos, Av. Unisinos 950, 93001-970
São Leopoldo, RS, Brasil *Instituto Nacional de Pesquisas da Amazônia,
Manaus, AM, Brasil
+ Corresponding author. Fax: +55-51-2590.8122. E-mail:strieder@cirrus.unisinos.br
This research received financial support from
the Universidade do Vale do Rio dos Sinos, Fapergs, and was partially supported
with a grant from MEC/Capes, Brazil, through the Programa Institucional de Aperfeiçoamento
de Pessoal de Nível Superior - PICDT, and is part of the first author's
PhD thesis.
Received 19 January 2001
Accepted 18 September 2001
Code Number: oc02011
The black flies of the genus Inaequalium
present a Neotropical distribution, with Panama at the northern limit, and the
Argentinian pampas at the southern, but do not occur in the Central Amazon.
This study offers a cladistic analysis establishing a hypothesis of relationships
between the species of Inaequalium. A total of 37 characters have been
considered in order to establish the hypothetic phylogenetic relationships.
Cerqueirellum (Py-Daniel, 1983) was considered as outgroup. Data were
analyzed using Henning 86 version 1.5. Wich the ie* command and implicit enumeration
a unique possible cladogram was obtained in Inaequalium with 52 steps,
and a CI of 0.76 and RI of 0.81. Two well-defined clades was obtained in the
resulting cladogram, the "botulibranchium" species-group, includes
I. travassosi, I. souzalopesi, I. botulibranchium and I. petropoliense,
and the "inaequale" species-group, includes I. rappae,
I. nahimi, I. inaequale, I. leopoldense, I. subnigrum, I. diversibranchium,
I. mariavulcanoae, I. nogueirai, I. beaupertuyi, I. clavibranchium and I.
subclavibranchium.
Key words: Neotropical - aquatic insects - black
flies - Simuliidae - Inaequalium
The geographic distribution of the genus Inaequalium
(Coscarón & Wygodzinsky, 1984) comprises South and Central America;
its northern limit is Panama and to the south it reaches the Pampas region in
Argentina, but is not found in Central Amazon. Most species are recorded from
the coastal mountains of Brazil (Coscarón & Wygodzinsky 1984, Coscarón
1987, 1991, Py-Daniel & Moreira 1989, Strieder et al. 1992, Coscarón
& Coscarón-Arias 1995, 1997, Strieder & Py-Daniel 1999, 2000).
Coscarón and Wygodzinsky established the
taxon Inaequalium in 1984 as a subgenus of Simulium Latreille,
1802, having Simulium (Inaequalium) inaequale (Paterson
& Shannon, 1927) as the type species. Adults of the eight species initially
included in Inaequalium, are black of medium size with wings between
2.3-2.8 mm long; thorax laterally and posteriorly with a gray stripe; cibarium
of females with an elevated base in the median area, with conspicuous, well
sclerotized spines. Females also present short paraprocts, much wider than long.
Males have basistyle with length and width almost equal and longer than the
dististyle; dististyle presents a sub-triangular, sub-quadrate or sub-cylindrical
shape and has strong apical or pre-apical spines. Pupae have cocoons with a
dorsal length between 2.1-3.8 mm and ventral length between 2.8-4.5 mm; filamentous
gills with six terminal filaments ("inaequale" species-group)
or thick gills with two to six branches ("botulibranchium"
species-group). Larvae present 1 + 1 sub-triangular posterior ventral tubercles;
generally deep gular cleft, with a sub-triangular shape or reduced cleft, with
a semicircular format (I. souzalopesi); the third article of the antenna in
general darker and longer than the first and second (Coscarón & Wygodzinsky
1984).
According to PyDaniel (1990) and PyDaniel and
Moreira Sampaio (1994, 1995), the recognition and utilization of the levels
of subfamily, tribe, genus and subgenus varies in the Simuliidae and that depends
on the point of view of the taxonomist. These authors, based on a revision of
Nearctic and Neotropical genera originally considered as Prosimuliini and the
genus Simulium from Simuliini, proposed to eliminate the tribal level
and to elevate all the subgenus of Simulium s.l. to generic rank. Under
these circumstances, Simulium Latreille, 1802 would be restricted to
Simulium s. str., in the Holarctic Zoogeographical region. Although many
taxonomists follows Crosskey and Howard (1997) which maintain the higher classification
of Crosskey (1987) and critique Py-Daniel and Moreira Sampaio (1994, 1995) hypothesis.
Coscarón et al. (1999) studied the species of Simulium from the
Neotropical region, after meticulous phylogenetic studies, these authors concluded
that the revised Neotropical species form an independent supraspecific group
of Simulium s. str., for which the name Asphatia was suggested.
In this work, the elevation of the subgenus to
genus in Simuliidae is followed, according to the nomenclature used by PyDaniel
and Moreira Sampaio (1994, 1995), Py-Daniel (1997), Strieder and Py-Daniel (1999,
2000) and Andrade et al. (2000). Since species of the genus Inaequalium
show a homogeneous adult morphology it is necessary to intensify the search
for new characters to facilitate recognition and in order to correlate the characters
of different stages. The larvae and pupae present features that allow the differentiation
of species (Coscarón & Wygodzinsky 1984, Strieder 1998, Strieder &
Py-Daniel 1999, 2000). The present study provides new characters for recognition
of species of Inaequalium and by a comparative analysis of these characters,
presents a phylogenetic hypothesis for the genus.
MATERIALS AND METHODS
Following Coscarón and Wygodzinsky (1984),
Crosskey (1987), Crosskey and Howard (1997), Coscarón (1987, 1991), Py-Daniel
and Moreira (1989), Py-Daniel and Moreira Sampaio (1995), Strieder and Py-Daniel
(1999, 2000) a preliminary list of the species of simuliids included in the
genus Inaequalium (Coscarón & Wygodzinsky, 1984) was prepared.
From the revision of types, the examination of material available in important
collections and bibliographic data a matrix of characters was elaborated. New
characters that allow a better differentiation of the species were verified.
Elaboration of the relationship between the species of Inaequalium was
based on the phylogenetic systematics developed by Henning (1966). On the table
that originated the data matrix of characters a code for each one was designed,
in which the number "0" (zero) means hypothetically the plesiomorphic
state, the numbers "1" (one) and "2" (two) indicate the
apomorphic state and "?" indicate that the character is not comparable
or unknown for the taxon. In the polarization of the characters considered for
the elaboration of a phylogenetic hypothesis among the species of Inaequalium,
the taxon Cerqueirellum (Py-Daniel, 1983), which correspond to a monophyletic
group nearly related, was considered as outgroup (Coscarón 1987). Data
were analyzed using the Tree Gardener 1.0 program (Ramos 1996), which was developed
based on Henning 86 version 1.5 (Farris 1989). The ie* command and implicit
enumeration were used for calculating the cladogram. For the majority of the
characters there was no distinction or emphasis. However, the character 12 was
considered particularly additive. The convergent characters (c) and the reversions
(r) are indicated on the cladogram obtained.
Only 15 species of the genus Inaequalium:
I. beaupertuyi (RamírezPérez, Rassi & Ramírez, 1977),
I. botulibranchium (Lutz, 1910), I. clavibranchium (Lutz, 1910),
I. diversibranchium (Lutz, 1910), I. inaequale (Paterson &
Shannon, 1927), I. leopoldense Strieder & Py-Daniel, 2000, I.
mariavulcanoae (Coscarón & Wygodzinsky, 1994), I. nahimi
(PyDaniel, 1984), I. nogueirai (D'Andretta & González, 1962),
I. petropoliense (Coscarón, 1980), I. rappae (PyDaniel &
Coscarón, 1982), I. subclavibran-chium (Lutz, 1910), I. subnigrum
(Lutz, 1910), I. souzalopesi (Coscarón, 1980) and I. travassosi
(D'Andretta & D'Andretta, 1947) were considered as final taxa in the phylogenetic
analysis. The others related species: I. clarki (Fairchild, 1940), I.
lurybayae (Smart, 1944), I. parimaensis (Ramírez Pérez,
Yarzábal, Takaoka, Tada & Ramírez, 1986), I. pseudoexiguum
(Nunes & Almeida, 1974), were not included due to their uncertain taxonomic
situation and/or insufficient data.
RESULTS AND DISCUSSION
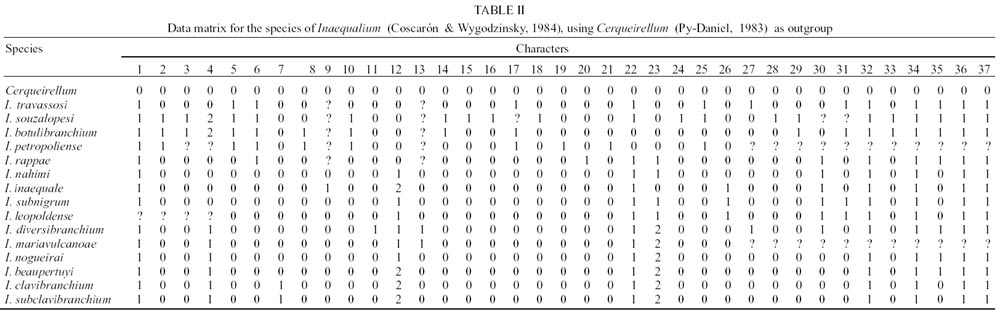
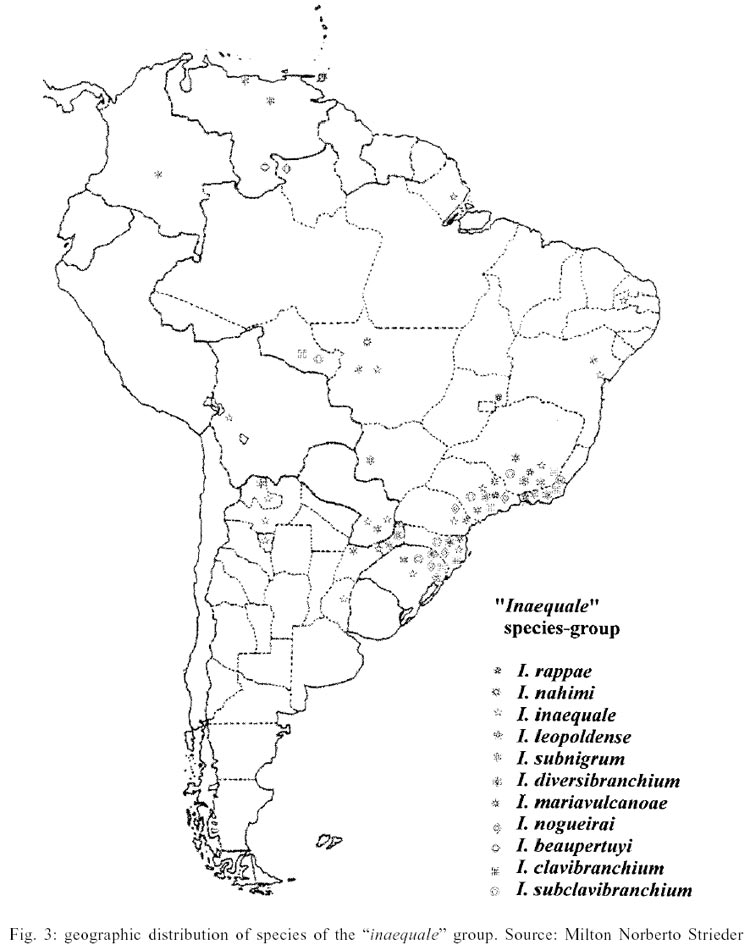
In the present phylogenetic study 37 characters
(2 for females, 2 for males, 22 for pupae and 11 for larvae) were used (Table
I a b).
Table II shows the basic matrix related
to the results of the analysis of the 37 characters considered. Using the program
Hennig 86, ie* command, a unique possible cladogram presenting data of the characters
considered was obtained. It presents a length of 52 steps, and a CI of 0.76
and RI of 0.81 (Fig. 1).
Cerqueirellum represents a monophyletic
taxon sustained by three synapomorphies derived from the morphology of the larva,
as follows: setae of cephalic apotome with internal branching; second external
teeth of the mandible absent or extremely reduced; and mandible with a row of
setae reaching the supramarginal inferior area. In the resulting cladogram of
Inaequalium, the species of the whole internal group are grouped independently
of Cerqueirellum, and this shows that these taxa are monophyletic.
The monophyly of the genus Inaequalium is
sustained by the following characters: basal portion of female cibarium with
small teeth arranged over median prominency (1:1); and third anterior part of
distal brush of larval mandible presenting setae with bifurcation near insertion
base (36:1). This synapomorphies observed to Inaequalium justify the
treatment of this group as genus, instead of subgenus. This is in agreement
with Coscarón and Wygodzinsky (1984), Coscarón (1987) and Coscarón
and Coscarón-Arias (1997), who concluded that Inaequalium (as a
subgenus) form a monophyletic taxon. The cladogram obtained in Inaequalium
has two well-defined clades, the "botulibranchium" species-group
and the "inaequale" species-group, similar to those of Coscarón
(1987) and Coscarón and Coscarón-Arias (1997).
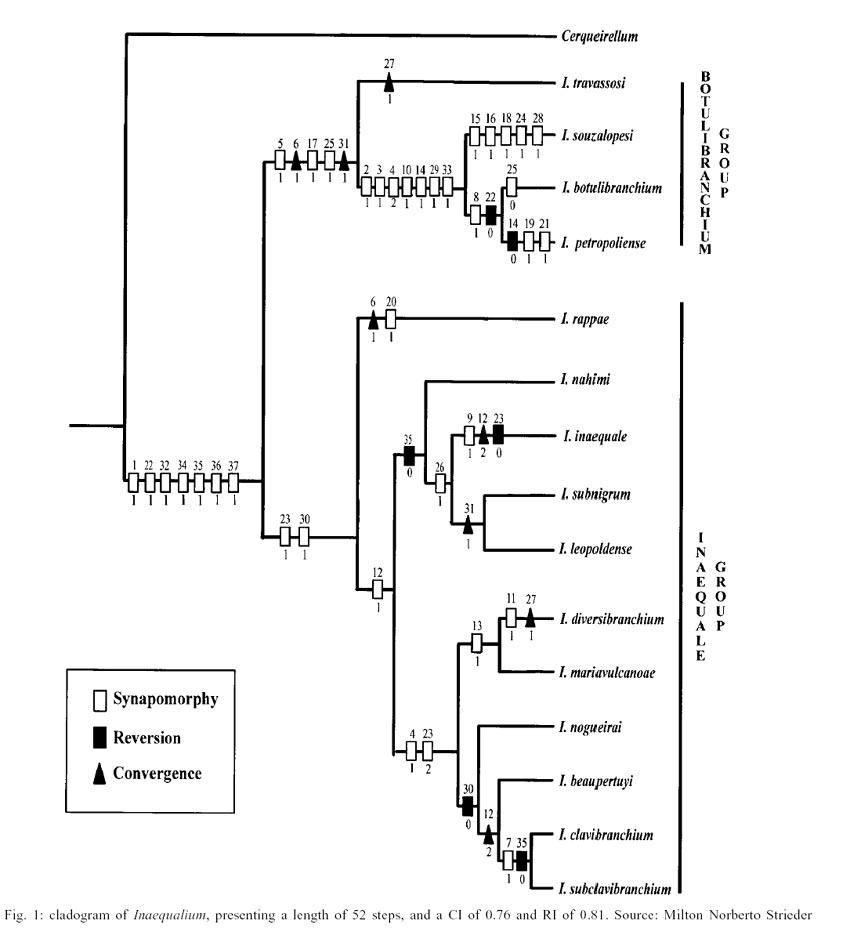
The "botulibranchium" species-group
includes I. travassosi, I. souzalopesi, I. botulibranchium and
I. petropoliense. This clade is defined by three synapo-morphies and
two convergences: (5:1) texture of pupal gill filaments without reticulate;
(6:1) pupae with thick gill branches (c); (17:1) pupal frontoclypeus with multinodulous
tubercles; (25:1) pupal abdomen tergites I and II with spines; and (31:0) larvae
without setae on dorsal-posterior region of abdominal cuticle (c). This group
has a distribution restricted to the mountains of southeastern Brazil (Fig.
2).
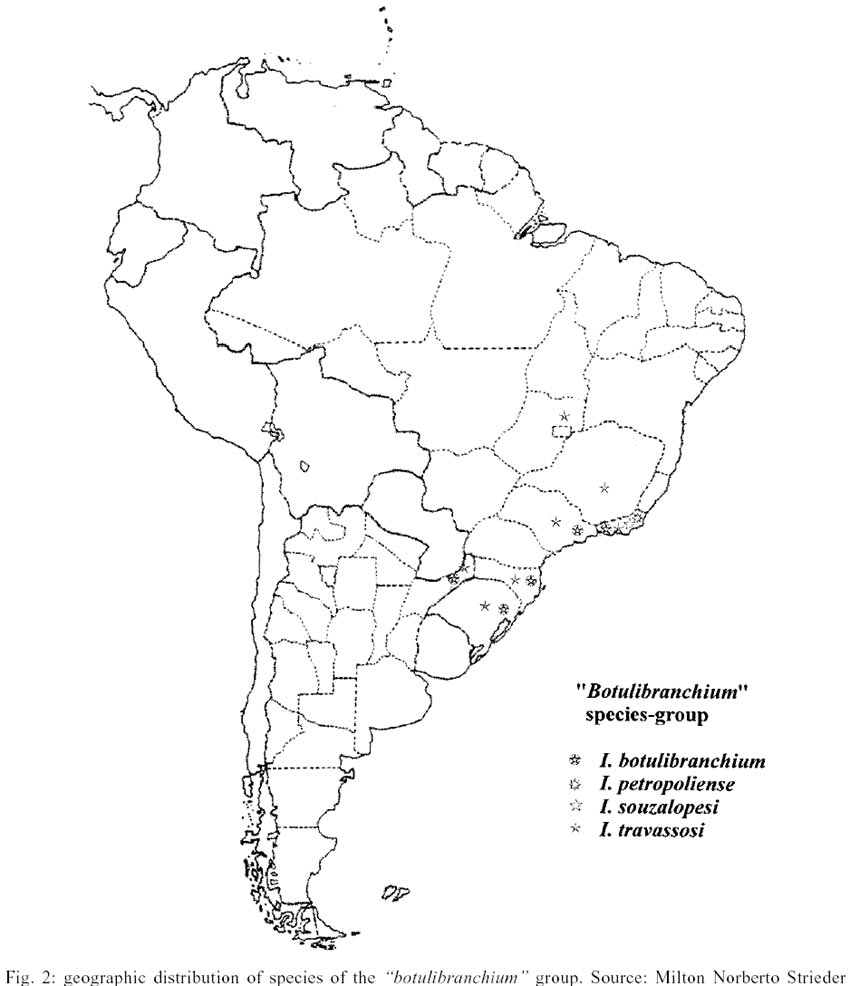
The "inaequale" species-group
includes I. rappae, I. nahimi, I. inaequale, I. leopoldense, I. subnigrum,
I. diversibranchium, I. mariavulcanoae, I. nogueirai, I. beaupertuyi, I. clavibranchium
and I. subclavibranchium. This group is defined by the following
synapomorphies: (23:1) and (23:2) pupa with thorax tubercles wider or as wide
as the diameter of the thoracic trichomes at point of insertion; (30:1) larval
anal gills with 13 to 20 lobes in each external branch.
This second species-group is also predominantly
from the mountains of southeastern Brazil with a larger diversity in the Serra
do Mar region, where only I. beaupertuyi and I. nahimi do not
occur (Fig. 3). The species I. inaequale
and I. subnigrum present a large range, covering most of the tropical
area of South America, except the lowland region of the Central Amazon. The
only two species that are absent from the mountains of southeastern Brazil occur
respectively on the Escudo Venezolano and in the southern region of Amazon,
restricted to the Mato Grosso in the old western Brazilian shield.
In relation to the two groups obtained in Inaequalium,
the results are similar to those presented by Coscarón (1987, 1991) and
Coscarón and Coscarón-Arias (1997). The results only disagree with
these authors in relation to the position of I. travassosi, which they
include in the "inaequale" group.
The first branching of the "botulibranchium"
species-group separates I. travassosi from I. souzalopesi subgroup.
I. travassosi present a homoplasy: (27:1) antenna of larva smaller than
the stem of labral fan (c).
I. souzalopesi subgroup present seven
synapo-morphies: (2:1) female paraproct basally longer than wide and acuminate
on distal half; (3:1) male distimere longer than twice its width at base; (4:2)
distimere with subcylindrical shape; (10:1) larval gill with ventral branch
curved; (14:1) pupal frontoclypeus and thorax with few tubercles; (29:1) larval
cervical sclerites big and fixed on cephalic apotome; and (33:1) larval hypostomial
bridge equal to or bigger than hypostomium.
The characters that separate I. souzalopesi
from I. botulibranchium and I. petropoliense are the following:
(15:1) and (16:1) pupal frontoclypeus with subdivided tubercles and without
nodulosity; (18:1) and (24:1) larva with simple cephalic and thoracic trichomes;
(28:1) antennal articles of larva with transversal striations.
I. botulibranchium subgroup is defined
by a synapo-morphy and a homoplasy: (8:1) pupal gill branches with appendages
(microfilaments or spines); (22:0) pupae with rounded and subtriangular tubercles
(r).
One character of the pupa, (25:0) abdominal tergites
I and II without spines (r), characterize I. botulibranchium.
I. petropoliense is characterized by one
homoplasy and two autoapomorphies of the pupa, as follows: (14:0) frontoclypeus
with abundant tubercles (r); (19:1) antennal sheaths with tubercles; and (21:1)
thoracic tubercles subtriangular shaped and with uniform distribution.
The first branching of the "inaequale"
group separates I. rappae from the other species.
I. rappae presents a homoplasy and an
autopomorphy: (6:1) pupal gill with thick branches (c); (20:1) pupal cephalic
genae with trichomes.
One synapomorphy and a homoplasic state: ventral
cocoon length almost equal (12:1) or smaller (12:2) than length of gill branches
supported the monophyly of the species with filiform gills.
The second branching of the "inaequale"
group results in two clades, respectivaly supported with a reversion and two
synapomorphies. These two monophyletic groups are composed by four and six species
respectively. The homoplasy: (35:0) lateral mandibular process straight or with
a slight depression in the middle area (r), is the character which congregates
the following species: I. nahimi, I. inaequale, I. leopoldense and
I. subnigrum. The synapomorphies: (4:1) distimere subquadrate shaped; and
(23:2) pupa with wide thoracic tubercles, relating to the diameter of thoracic
trichomes at point of insertion, sustain the monophyly of the following species:
I. diversibranchium, I. mariavulcanoae, I. nogueirai, I. beaupertuyi,
I. clavibranchium and I. subclavibranchium.
The first branching of the first clade of species
with filiform gills separates I. nahimi from I. inaequale subgroup.
I. inaequale subgroup is defined for:
(26:1) presenting tubercles in the abdomen tergite I of the pupa. The next branching
separates I. inaequale from I. subnigrum and I. leopoldense.
I. inaequale presents a synapomorphy and
two homoplasies: (9:1) filiform gills with the primary ventral branch and the
two secondary dorsal branches generally of the same length; (12:2) cocoon length
smaller than the maximum length of the gill filaments (c); and (23:0) pupal
thoracic tubercles small, relative to the diameter of the thoracic trichomes
at the point of insertion (r).
I. subnigrum and I. leopoldense
present a homoplasy: (31:1) setae from abdominal cuticle on the dorsal-posterior
region of larva absent or in a reduced number (c).
The first branching of the second clade of species
with filiform gills separates I. diversibranchium and I. mariavulcanoae
from I. nogueirai subgroup.
I. diversibranchium and I. mariavulcanoae
present a synapomorphy: (13:1) filiform gills with the primary dorsal branch
presenting bigger or equal length to ventral primary branch.
The next dichotomy separates I. nogueirai
from I. beaupertuyi subgroup.
I. beaupertuyi subgroup presents a homoplasy:
(12:2) ventral length of cocoon smaller than the maximum length of the gill
filament (c).
I. clavibranchium and I. subclavibranchium
establish a monophyletic group, apart from I. beaupertuyi by a synapomorphy
and a homoplasy: (7:1) gill filaments of pupa with distal portion inflated;
(35:0) lateral mandibular process almost straight and not crossing inferior
margin of mandible (r).
This work presents a cladistic analysis of the
genus Inaequalium (Coscarón & Wygodzinsky, 1984), which allows
the following conclusions to be made: (1) the analysis of 37 characters confirms
the existence of two clades morphologically well defined in this taxon. The
"botulibranchium" group, which brings together the species:
I. travassosi, I. souzalopesi, I. botulibranchium and I. petropoliense;
and the "inaequale" group, which brings together the following
species: I. rappae, I. nahimi, I. inaequale, I. subnigrum, I. leopoldense,
I. diversibran-chium, I. mariavulcanoae, I. nogueirai, I. beaupertuyi, I.
clavibranchium and I. subclavibranchium; (2) Inaequalium is
a monophyletic group, which presents a wide geographic distribution in the Neotropical
region occurring predominantly in the coastal region of Brazil, with a larger
diversity on the slopes of the Serra do Mar mountain range.
ACKNOWLEDGEMENTS
To Dr Sixto Coscarón, Universidad Nacional
de La Plata, Museo de La Plata, Argentina, for the incentive and for lending
specimens; to Dr Cecília Volkmer-Ribeiro, Fundação Zoobotânica
do Rio Grande do Sul; Dr Gervásio Silva Carvalho, Pontifícia Universidade
Católica do Rio Grande do Sul; Dr Gilson Moreira, Universidade Federal
do Rio Grande do Sul for helpful discussions. To Dr Paulo Günter Windisch,
Fernando Joner and José Eloy dos Santos Jr, Universidade do Vale do Rio
dos Sinos, for reviewing and improving the manuscript.
REFERENCES
- Andrade HTA, Nascimento RSS, Gurgel HCB, Medeiros
JF 2000. Simuliidae (Diptera) integrantes da dieta de Poecelia vivipara
Bloch & Schneider, 1901 (Atheriniformes; Poeciliidae) no rio Ceará-Mirim,
Estado do Rio Grande do Norte, Brasil. Entomol y Vec 7: 119-122.
- Coscarón S 1987. El gênero Simulium
Latreille en la Región Neotropical: Análisis de los Grupos Supraespecíficos,
Especies que los Integran y Distribución Geográfica (Simuliidae,
Diptera), Museu Paraense Emílio Goeldi, Coleção Emilie
Snethlage, Belém, Brazil, 111 pp.
- Coscarón S 1991. Simuliidae. Fauna
de Agua Dulce de la República Argentina (Insecta, Diptera), Fundación
para la Educación, la Ciencia y la Cultura, Buenos Aires, 304 pp.
- Coscarón S, Coscarón-Arias CL 1995.
Distribution of Neotropical Simuliidae (Insecta, Diptera) and its areas of
endemism. Rev Acad Colomb Cienc 19: 717-732.
- Coscarón S, Coscarón-Arias CL 1997.
Cladistic analysis of the subgenera Inaequalium, Psaroniocompsa,
and Chirostilbia of the genus Simulium, with comments on their
distribution (Diptera: Simuliidae). Rev Soc Entom Arg 56: 109-121.
- Coscarón S, Wygodzinsky P 1984. Notas
sobre simúlidos neotropicales. VII. Sobre los subgêneros Psaroniocompsa
Enderlein y Inaequalium, subgen. nov. Arq Zool São Paulo
31: 37103.
- Coscarón S, Ibáñez-Bernal S,
Coscarón-Arias CL 1999. Revision of Simulium (Simulium)
in the Neotropical realm (Insecta: Diptera: Simuliidae). Mem Entomol Internl
14: 543-604.
- Crosskey RW 1987. An annotated checklist of
the world black flies (Diptera: Simuliidae), Part. VIII, Black fly species
of the world. In KC Kim, RW Merritt (eds), Black Flies: Ecology, Population
Management, and Annotated World List, University Park: The Pennsylvania
State University, p. 425-520.
- Crosskey RW, Howard TM 1997. A New Taxonomic
and Geographical Inventory of World Blackflies (Diptera: Simuliidae),
The Natural History Museum, London, 144 pp.
- Farris JS 1989. HENNIG86. A PC-DOS
Program for Phylogenetic Analysis. Cladistics 5: 1-163.
- Hennig W 1966. Phylogenetic Systematics,
University of Illinois Press, Urbana, 263 pp.
- Py-Daniel V 1983. Caracterização
de dois novos subgêneros em Simuliidae (Diptera: Culicomorpha) Neotropical.
Amazoniana 8: 159-223.
- Py-Daniel V 1990. Revisão da Tribo
Prosimuliini (Diptera: Culicomorpha), Simuliidae nas Regiões Zoogeográficas
Neártica e Neotropical (Larvas e Pupas), PhD Thesis, Instituto de
Pesquisas da Amazônia, Manaus, 334 pp.
- Py-Daniel V 1997. Oncocercose, uma endemia
focal no hemisfério norte da Amazônia. In RI Barbosa, EJG Ferreira,
EG Castellón (eds), Homem, Ambiente e Ecologia no Estado de Roraima,
Inpa, Manaus, p. 111-155.
- Py-Daniel V, Moreira GRP 1989. Simuliidae
(Diptera, Culicomorpha) no Brasil. IX. Sobre o Simulium (Inaequalium) nogueirai
d'Andretta & González, 1964. Iheringia 69: 85-95.
- Py-Daniel V, Moreira Sampaio RT 1994. Jalacimgomyia
gen.n. (Culicomorpha); a ressurreição de Gymnopaidinae; a eliminação
do nível tribal; apresentação de novos caracteres e a redescrição
dos estágios larval e pupal de Simulium colombaschense (Fabricius,
1787) (Diptera: Simuliidae). Mem CAICET 4: 101-148.
- Py-Daniel V, Moreira Sampaio RT 1995. Gêneros
e espécies de Simuliidae (Diptera: Culicomorpha) assinalados para o Brasil
até 1995. Entomol y Vec 2: 117-121.
- Ramos TC 1996. Tree Gardener version 1.0 software
implementation. Distributed by the author.
- Strieder MN 1998. Revisão e Análise
Cladística de Espécies de Inaequalium (Coscarón & Wygodzinsky,
1984) (Diptera, Simuliidae), PhD Thesis, Pontifícia Universidade
Católica do Rio Grande do Sul, Porto Alegre, 160 pp.
- Strieder MN, Py-Daniel V 1999. Espécies
de Inaequalium (Diptera, Simuliidae): dados bionômicos e chaves
para sua identificação. Biociências 7: 43-72.
- Strieder MN, Py-Daniel V 2000. Revisão
de Inaequalium (Diptera, Simuliidae), com redescrição das
formas imaturas e descrição de uma nova espécie. Entomol
y Vec 7 (Suppl. 2): 1-91.
- Strieder MN, Corseuil E, Py-Daniel V 1992.
Espécies do gênero Simulium (Diptera, Simuliidae) ocorrentes
no Rio Grande do Sul, Brasil, com chaves para sua identificação.
Acta Biol Leopold 14: 53-74.
© 2002
Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02011t1a.jpg]
[oc02011t2.jpg]
[oc02011t1b.jpg]
[oc02011f2.jpg]
[oc02011f3.jpg]
[oc02011f1.jpg]
| 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}