
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 1, 2002, pp. 95-99
|
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(1) 2002, pp. 95-99
Additives and Protein-DNA Combinations
Modulate the Humoral Immune Response Elicited by a Hepatitis C Virus Core-encoding
Plasmid in Mice
Liz Alvarez-Lajonchere/+, Santiago
Dueñas-Carrera, Ariel Viña, Thelvia Ramos*, Dagmara Pichardo,
Juan Morales
HCV Department, Vaccine Division, Centro de Ingeniería
Genética y Biotecnología *Centro Nacional de Genética Médica,
Casilla Postal 6162, Havana City, Cuba
+Corresponding author. Fax: +53-7-214764. E-mail:
juan.morales@cigb.edu.cu
Received 20 February 2001
Accepted 10 October 2001
Code Number: oc02017
Humoral and cellular immune responses are
currently induced against hepatitis C virus (HCV) core following vaccination
with core-encoding plasmids. However, the anti-core antibody response is frequently
weak or transient. In this paper, we evaluated the effect of different additives
and DNA-protein combinations on the anti-core antibody response. BALB/c mice
were intramuscularly injected with an expression plasmid (pIDKCo), encoding
a C-terminal truncated variant of the HCV core protein, alone or combined with
CaCl2, PEG 6000, Freund's adjuvant, sonicated calf thymus DNA and
a recombinant core protein (Co.120). Mixture of pIDKCo with PEG 6000 and Freund's
adjuvant accelerated the development of the anti-core Ab response. Combination
with PEG 6000 also induced a bias to IgG2a subclass predominance among anti-core
antibodies. The kinetics, IgG2a/IgG1 ratio and epitope specificity of the anti-core
antibody response elicited by Co.120 alone or combined with pIDKCo was different
regarding that induced by the pIDKCo alone. Our data indicate that the antibody
response induced following DNA immunization can be modified by formulation strategies.
Key words: hepatitis C virus - DNA immunization
- core
It is now well established that injection of
plasmid DNA through a wide range of routes induces both humoral and cellular
immune responses against the encoded proteins in several hosts. Moreover, immune
responses induced by DNA immunization have lead to protection against various
viral, bacterial and parasitic pathogens (Donnelly et al. 1997). However, immune
responses generated by this methodology against several antigens remain insufficient
(Hasan et al. 1999). While the majority of the experiments conducted to date
have used phosphate buffered saline pH 7.5 (PBS) or saline as a diluent for
the injected DNA, a great deal of effort is now being applied to the development
of delivery vehicles and adjuvants. Such reagents may increase the uptake of
DNA, reduce the necessary dose for immunization and enhance subsequent immune
responses. Some systems currently under investigation are cationic liposomes
(Gregoriadis et al. 1997), immuno-stimulatory oligonucleotide sequences (Klinman
et al. 1997), cytokines (Chow et al. 1997) and monophosphoryl lipid A (Sasaki
et al. 1998).
In this paper, different formulations were investigated
to modulate the antibody response generated by a DNA vaccine. We studied the
humoral immune response elicited by vaccination with a plasmid encoding a truncated
variant of the hepatitis C virus (HCV) core protein. Several B cell and cytotoxic
T-lymphocyte determinants within the HCV core protein have been characterized
(Walker 1996, Jackson et al. 1997). Moreover, of the various regions of HCV,
antibodies against the core protein are often the first to appear during HCV
natural infection (Okamoto et al. 1992). However, plasmid expressing the HCV
nucleocapsid alone often elicited strong cellular but weak and/or transient
humoral immunity (Lagging et al. 1995, Chen et al. 1995). Consequently, the
HCV core antigen is a good model for investigations about the modulation of
antibody response generated following DNA immunization. We particularly investigated
different additives to facilitate the plasmid stability and uptake by the cells.
Use of a classical adjuvant like Freund and the combination of plasmid DNA with
a recombinant core protein were also evaluated to obtain an improved anti-core
antibody response.
MATERIALS AND METHODS
Co.120 protein - Recombinant Co.120 is
an Escherichia coli-derived protein containing the first 120 amino acids
of HCV viral polyprotein. It was purified by a combination of washed pellet
procedures and gel filtration chromatography as previously described (Dueñas-Carrera
et al. 1999). Briefly, recombinant protein was expressed in BL21 (DE3) E.
coli cells. For purification, expressing cells were disrupted with French
press (Braund, 1,500 kg/cm2) at 1 g/ml in 10 mM Tris HCl, 6 mM EDTA,
pH 8. The insoluble fraction of cell lysate was washed with 0.5 M urea, 1% Triton
X-100, 10 mM EDTA, 300 mM NaCl, 10 mM 2-mercaptoethanol, 10 mM Tris HCl pH 8
buffer. Co.120 protein was solubilized by increasing the concentration of the
urea up to 2 M. The supernatant was desalted on a Sephadex-G25 column equilibrated
with carbonate buffer, pH 10.6. Desalting by gel filtration rendered soluble
Co.120 protein at 95% of purity.
Human sera - Human sera were obtained
from blood donors and chronic patients and previously screened for the presence
of anti-HCV antibodies by UMELISA HCV from Centro de Inmunoensayo (La Habana,
Cuba). The anti-HCV positive sera were also confirmed by Ortho HCV 2.0 ELISA
(Ortho Diagnostic Systems, Raritan, NJ).
Plasmids - pAEC-K6: expression vector
that contains the human cytomegalovirus immediate early promoter, simian virus
40 terminator and polyadenylation sequences, a bacterial replication origin
from pUC and a kanamycin resistant gene as selection marker. pIDKCo: an expression
plasmid generated by inserting a 528 nucleotide DNA fragment, coding for the
first 176 aa of the HCV core protein, into the compatible sites of the pAEC-K6
(Dueñas-Carrera et al. 2000).
E. coli strain XL-1 Blue [(F', proAB,
lacIqDZM15, Tn10), endA, hsdR17, supE,
thi-1, recA1, gyrA96, relA1, lac.] was transformed with pIDKCo and pAEC-K6 plasmids.
Cells were grown under selective pressure with 50 mg/l kanamycin. Plasmid DNA
was subsequently purified as previously described (Horn et al. 1995).
Animals and immunization schedule - BALB/c
female mice of 6 to 8 weeks old (18-20 g of weight) were purchased from CENPALAB
(Ciudad de la Habana, Cuba) and hosted in appropriated animal care facilities
during the experimental period. The animals were handed following the international
guidelines required for experimentation with animals. Mice were injected at
0 and 3 weeks, in the quadriceps muscle, with 100 µl of different immunogens.
Blood samples were collected from the retro-orbital sinus at 0, 5, and 14 weeks
after the primary immunization. The mice were euthanized after the final blood
samples were taken. The study included 10 groups of 5 mice each. The groups
were conformed as follows: group 1 (pAEC-K6) was injected with a mock DNA (pAEC-K6
plasmid). The second group (Co.120-pIDKCo) received pIDKCo combined with the
protein Co.120. Both components were blended and stirred overnight. This mixture
was centrifuged for 15 min at 3,000 xg and the supernatant was given as immunogen.
The third (Co.120) and fourth (pIDKCo) groups received the Co.120 protein and
the pIDKCo alone, respectively. Groups 5 (CaCl2), 6 (PEG 6000), 7
(Freund's) and 8 (sCT-DNA) were immunized with pIDKCo combined with 100 mM CaCl2
(Merck, Darmstadt, Germany), 1% PEG 6000 (Merck, Darmstadt, Germany), Freund's
adjuvant (Sigma, St Louis, USA) and 100 µg of sonicated calf thymus DNA
(sCT-DNA) (Promega, Madison, USA), respectively. Group 9 (pIDKCo/Co.120) was
primed with pIDKCo and boosted with Co.120. The last group (Co.120/pIDKCo) was
primed with Co.120 and boosted with pIDKCo. Plasmids were administered at 1
µg/µl and Co.120 protein at 0.1 µg/µl in PBS.
Enzyme-linked immunosorbent assay (ELISA)
- To detect murine anti-core antibodies, 96-well microtiter plates (Costar,
Cambridge, MA, USA) were coated with 100 µl of Co.120 (10 µg/ml)
diluted in coating buffer (50 mM carbonate buffer, pH 9.6) overnight at 4ºC.
The wells were washed three times with 0.05% Tween 20 in PBS (PBST) and blocked
with 200 µl of PBST containing 1% skimmed milk (Oxoid, Basingstoke, Hampshire,
England) for 1 h at room temperature. After three washes with PBST, 100 µl
of serial two-fold dilutions of individual mouse sera in PBST were added and
incubated at 37ºC for 1 h. The plates were washed three times with PBST,
and 100 µl of horseradish peroxidase-conjugate goat anti-mouse IgG (Amersham,
Little Chalfont, Bucks, UK) 1:3000 diluted was added at 37ºC for 1 h. For
subtyping of mouse antibodies, 1:50 dilution of pooled sera from each group
was used. Biotinylated anti-mouse IgG of the appropriate subclass (Amersham,
Little Chalfont, Bucks, UK) was added at 1:1000 dilution, followed by a streptavidin-biotinylated
horseradish peroxidase conjugated (Amersham, Little Chalfont, Bucks, UK) diluted
1:3000 and incubated for 30 min at 37ºC. Positive reactions were visualized
with o-phenylenediamine (Sigma, St Louis, USA) in 0.1 M citric acid, 0.2 M,
NaH2PO4, pH 5.0 and 0.015% H2O2
as substrate; the reaction was stopped with 50 µl of 2.5 M, H2SO4.
Measurement of optical density (OD) at 492 nm was made in a plate reader (Merck,
Darmstadt, Germany).
The cut-off value to consider a positive mouse
anti-core antibody response was established as twice the mean absorbance of
the pre-immune sera.
To determine if human anti-HCV positive sera
were able to bind to Co.120 protein, a similar ELISA was carried out. Human
sera were individually evaluated at 1:10 dilution in PBST. Anti-human IgG-peroxidase
conjugate (Amersham, Little Chalfont, UK), diluted 1:3000, was employed instead
of anti mouse IgG-peroxidase conjugate. The other steps were performed as described
above. The cut-off value employed to consider a positive human anti-core antibody
response was established as twice the mean absorbance of the negative control
human sera.
The competition of antibodies from pIDKCo plasmid-immunized
mice (PIM) with human anti-HCV positive sera for the binding to Co.120 protein
was also studied by using essentially the same ELISA. After blocking, human
sera at 1:10 dilution were added and incubated at 37ºC. One hour later,
after three washes with PBST, the plates were incubated for 1 h at 37ºC
with pooled sera from each group of PIM at 1:50 dilution. Anti mouse IgG-peroxidase
conjugate was also used at 1:3000 dilution. The inhibitory activity was expressed
as percentage of inhibition and determined as follows:
Percentage of inhibition = 100 x (A492nm
of PIM Ab – A492nm of PIM Ab bound to Co.120 in the presence
of human anti-HCV positive sera)/ A492nm of PIM Ab).
Statistical analysis - To compare differences
among groups, a One-way ANOVA with the Newman-Keuls post test was used. P
values < 0.05 were considered significant.
RESULTS
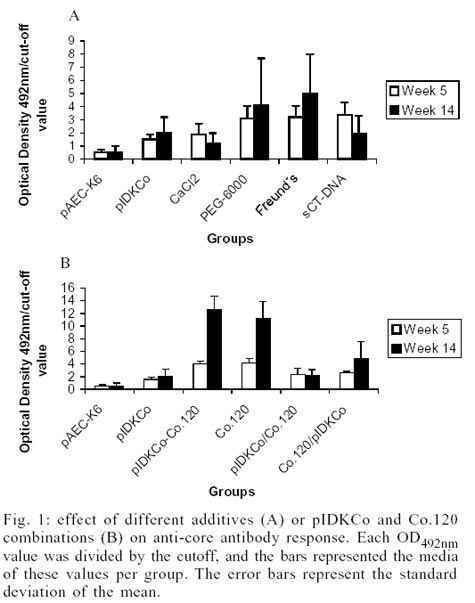
Influence of additives in the anti-core antibody
response - BALB/c mice were immunized with the pIDKCo plasmid alone or mixed
with different additives. Animals were observed through 14 weeks for the development
of anti-HCV core Ab response. Animals vaccinated with pAEC-K6 (control plasmid)
did not show any reactivity against Co.120 protein when compared with their
own preimmune sera. In contrast, five weeks after primary immunization, anti-core
antibodies were detected in 80 and 100% of mice vaccinated with pIDKCo alone
or mixed with the additives, respectively (data not shown).
The anti-capsid antibody response at week 5 and
14, determined as the OD492nm/cut-off ratio when the sera were diluted
1:50, is shown in Fig. 1A. The inclusion
of CaCl2 in the immunogen did not improve the humoral immune response.
On the other hand, the mixture of pIDKCo with PEG 6000, Freund's adjuvant and
sCT-DNA induced a statistically stronger anti-core Ab response than immunization
with pIDKCo alone (p < 0.01) at week 5 of the schedule. However, at week
14 there were not differences in the anti-core antibody response among the groups
of immunized mice.
Effect of different combinations of pIDKCo
with Co.120 on anti-core antibody response - To investigate the effects
of protein-DNA combinations on the anti-core Ab response, BALB/c mice were immunized
with pIDKCo plasmid and Co.120 protein in different formulations. Fig.
1B shows the anti-core antibody response determined as OD492nm/cut-off
ratio when the sera were diluted 1:50.
Five weeks after the primary immunization, all
groups of immunized mice had anti-core antibodies except the group vaccinated
with pAEC-K6. The combination of pIDKCo with Co.120 always produced a similar
anti-core Ab response than the protein alone, and in both groups the anti-core
antibody response was statistically higher than for the pIDKCo alone or the
prime-boost schedules (p < 0.05). There were no differences between the anti-core
antibody responses induced by prime/boost schedules. However, five weeks after
primary immunization, the priming with Co.120 and boost with pIDKCo rendered
statistically higher response than the group immunized with pIDKCo alone (p
< 0.01).
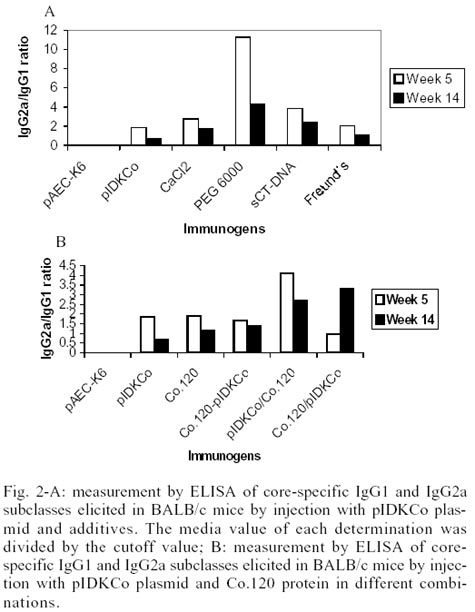
Characterization of anti-core antibody elicited
by the combination of plasmid with additives or protein - Sera from mice
immunized with pIDKCo or Co.120 protein showed a mixed pattern of IgG2a/IgG1
anti-core antibodies. However, a tendency for predominance of IgG2a Abs was
detected in the groups immunized with prime/boost schedules or pIDKCo-PEG 6000
and sCT-DNA combinations (Fig. 2A-B).
Furthermore, in order to characterize the specificity
of anti-core Ab response in mice, an inhibition of binding ELISA was carried
out (Table). Human sera were incubated
with Co.120 protein before incubation of mouse sera with this recombinant protein.
Results corresponding to sera from mice immunized with the mixture of pIDKCo
with CaCl2 were not analyzed because both positive and negative anti-HCV
human sera completely abolished the weak reactivity of these mouse sera to the
Co.120 protein. Positive anti-HCV human sera maximally inhibited the binding
of anti-core Ab induced by pIDKCo alone. Interestingly, our results showed that
the inclusion of additives and the combinations of plasmid with protein changed
the epitope specificity of anti-core antibodies.
DISCUSSION
DNA-based immunization appears promising as a
new way to administer vaccines. Such a vaccination strategy offers a means of
mimicking important characteristics of live attenuated viral vaccines like the
synthesis of native antigens and the induction of class I major histocompatibility
complex-restricted cytotoxic T lymphocyte responses. However, nucleic acid vaccines
do not seem to induce a response as strong as do conventional (lived attenuated)
vaccines and consequently different approaches have been used to modulate the
plasmid DNA immune responses. These efforts have been directed mainly to recruit
immune cells and to facilitate the entry of plasmid DNA to cells.
Previous studies have shown that the intramuscular
injection of plasmid expressing the HCV core protein was capable of inducing
detectable core-specific antibody response, lymphoproliferative responses and
cytotoxic T-lymphocyte activity in mice (Major et al. 1995).
In earlier attempts several doses of 100-200
µg of HCV core-encoding constructs have been generally administered in
mice every second week. Nevertheless, only weak humoral immune responses have
been induced against non-fused core variants by this way (Lagging et al. 1995,
Chen et al 1995, Hu et al. 1999). In contrast, we have previously demonstrated
that the pIDKCo plasmid induced a potent immune response following a DNA immunization
schedule of five doses with increasing intervals between them (Dueñas-Carrera
et al. 2000).
PEG has been used to increase the efficiency
of yeast transfection (Beggs 1978). Besides, this molecule has been used in
combination with cationic lipids to increase DNA transfection in mammalian cells
(Harvie et al. 2000). In fact, immunization with the combination of pIDKCo and
PEG 6000 produced a temporary augment of the anti-core antibody response. This
mixture also induced a bias through IgG2a subtype, suggesting the activation
of cellular branch of immune response. Transfection efficiency experiments are
required to correlate these effects with an increased plasmid uptake in vivo.
Since one of the problems of DNA immunization
is the degradation of plasmid by extracellular nucleases, a way to ensure cells
transfection is to protect the DNA from these enzymes. With this purpose we
employed the sonicated calf thymus DNA as a substrate competitor to diminish
nuclease specific degradation of the plasmid vaccine. This is the first report
in which sonicated calf thymus is employed with the idea of protecting plasmid
DNA from nucleases in vivo. Actually, we can not say that this molecule succeeded
in that function, but it certainly caused a transient increase in anti-core
antibody response when was combined with the pIDKCo plasmid. Remarkably, Weiner
et al. (1997) have demonstrated that calf thymus DNA, in contrast
to prokaryotic DNA, does not have an intrinsic adjuvant effect.
On the other hand, calcium salts have been used
by Wang et al. (2000) to increase antibody titers in a formulation of a DNA
vaccine encoding hepatitis B surface antigen. However, mixture of CaCl2
with pIDKCo did not stimulate the anti-core antibody response.
Prime-boost strategies have also been evaluated
to increase immune response to the HCV core antigen. Chen et al. (1995) demonstrated
that a second inoculation with the protein encoded by the vaccination plasmid
recruited lymphocyte clones primed by the first inoculation with DNA. In the
present work, only the prime Co.120/boost pIDKCo schedule transiently increased
the anti-core antibody response with respect to the generated after immunization
with pIDKCo alone. Moreover, the Co.120 protein alone or combined with the plasmid
pIDKCo induced higher anti-core antibody response than pIDKCo alone or boosted
with protein and vice versa. These results are consistent with the immunogenic
character demonstrated by Co.120 in mice and rabbits (Dueñas-Carrera
et al. 1999, Alvarez-Obregón et al. 2001), in spite of controversial
results regarding the immunogenicity of truncated nucleocapsid proteins in mice
(Inchauspe et al. 1997, Harase et al. 1997).
We compared the epitope specificities of anti-core
antibodies in PIM with those in HCV-immune sera by an inhibitory assay. The
information obtained in this experiment denoted that the human HCV-immune sera
had antibodies that could bind to the same regions in the HCV capsid that were
recognized by antibodies in mice immunized with plasmid DNA. The level of inhibitory
activity was not related with the reactivity of anti-HCV positive human sera
to the capture antigen.
Interestingly, sera from mice immunized with
plasmid combined with additives or Co.120 protein showed different behavior
in the inhibitory assay regarding those from mice immunized with pIDKCo alone.
Sera from mice immunized with pIDKCo evidenced the greatest inhibitory activity,
confirming that anti-core antibodies induced in mice by immunization with pIDKCo
were similar to those in human sera with respect to the epitope specificity
(Dueñas-Carrera et al. 2000).
The results presented here confirmed that vaccination
with HCV-core derived DNA sequences could be an effective method to induce humoral
response to HCV. Moreover, our data indicate that kinetics, magnitude and epitope
specificity of the anti-HCV core humoral immune response generated following
DNA immunization could be modified by formulation and prime/boost strategies.
ACKNOWLEDGEMENTS
To Dr Antonieta Herrera and Dr Alejandro Martin
for critical and constructive reading of the manuscript.
REFERENCES
- Alvarez-Obregon JC, Dueñas-Carrera
S, Valenzuela C, Morales J 2001. A truncated HCV core protein elicits a potent
immune response with a strong participation of cellular immunity components
in mice. Vaccine 19: 3940-3946.
- Beggs JD 1978. Transformation of yeast by
a replicating hybrid plasmid. Nature 275: 104-109.
- Chen Z, Wang RY-H, Alter HJ, Shih JW-K 1995.
Genetic immunization of mice with plasmid containing hepatitis C virus core
protein-encoding DNA. Vaccine Res 4: 135-144.
- Chow Y-H, Huang WL, Chi W-K, Chu Y-D, Tao
M-H 1997. Improvement of hepatitis B virus DNA vaccines by plasmids co-expressing
hepatitis B surface antigen and Interleukin-2. J Virol 71: 169-178.
- Donnelly JJ, Ulmer JB, Shiver JW, Liu MA 1997.
DNA vaccines. Annu Rev Immunol 15: 617-648.
- Dueñas-Carrera S, Alvarez-Lajonchere
L, Alvarez-Obregón JC, Herrera A, Lorenzo LJ, Pichardo D, Morales J
2000. A truncated variant of the hepatitis C virus core induces a slow but
potent response in mice following DNA immunization. Vaccine 19: 992-997.
- Dueñas-Carrera S, Morales J, Acosta-Rivero
N, Lorenzo LJ, García C, Ramos T, Guerra I, Peña M 1999. Variable
level expression of hepatitis C virus core protein in a prokaryotic system.
Analysis of the humoral response in rabbit. Biotecnología Aplicada
16: 226-231.
- Gregoriadis G, Saffie R, de Souza B 1997.
Liposome mediated DNA vaccination. FEBS Lett 402: 107-110.
- Harase I, Moriyama T, Kaneko T, Kita H, Nomura
M, Suzuki G, Ohnishi H, Muto Y, Yasaki Y, Imawari M 1995. Immune response
to hepatitis C virus core protein in mice. Immunol Cell Biol 73: 346-352.
- Harvie P, Wong FM, Bally MB 2000. Use of poly
(ethylene glycol)-lipid conjugates to regulate the surface attributes and
transfection activity of lipid-DNA particles. J Pharm Sci 89: 652-663.
- Hasan UA, Abai AM, Harper DR, Wren BW, Morrow
WJW 1999. Nucleic acid immunization: Concepts and techniques associated with
third generation vaccines. J Immunol Meth 229: 1-22.
- Horn NA, Meek JA, Budahazi G, Marquet M 1995.
Cancer gene therapy using plasmid DNA: purification of DNA for human clinical
trials. H Gene Ther 6: 565-573.
- Hu G-J, Wang RY-H, Han DS, Alter HJ, Shih
WK 1999. Characterization of the humoral and cellular immune responses against
hepatitis C virus core induced by DNA-based immunization. Vaccine 17:
3160-3170.
- Inchauspe G, Vitvitski L, Major ME, Jung G,
Spengler U, Maisonnas M, Trepo C 1997. Plasmid DNA expressing a secreted or
a nonsecreted form of hepatitis C virus nucleocapsid: comparative studies
of antibody and T-helper responses following genetic immunization. DNA
and Cell Biol 16: 185-195.
- Jackson P, Petrik J, Alexander GJM, Pearson
G, Allain J-P 1997. Reactivity of synthetic peptides representing selected
regions of hepatitis C virus core and envelope proteins with a panel of hepatitis
C virus-seropositive human plasma. J Med Virol 51: 67-79.
- Klinman DM, Yamschikov G, Ishigatsubo Y 1997.
Contribution of CpG motifs to the immunogenicity of DNA vaccines. J Immunol
158: 3635-3639.
- Lagging LM, Meyer K, Hoft D, Houghton M, Belshe
RB, Ray R 1995. Immune responses to plasmid DNA encoding the hepatitis C virus
core protein. J Virol 69: 5859-5863.
- Major ME, Vitvitski L, Mink MA, Schleef M,
Whalen RG, Trepo C, Inchauspe G 1995. DNA-base immunization with chimeric
vectors for the induction of immune responses against the hepatitis C virus
nucleocapsid. J Virol 65: 5798-5805.
- Okamoto H, Tsuda F, Machida A, Munekata E,
Akahane Y, Sugai Y, Mashiko K, Mitsui T, Tanaka T, Miyakawa Y, Mayumi M 1992.
Antibodies against synthetic oligopeptides deduced from the putative core
gene for the diagnosis of hepatitis C virus infection. Hepatology 15:
180-186.
- Sasaki S, Hamajima K, Fukushima J, Ihata A,
Ishii N, Gorai I, Hirahara F, Mohri H, Okuda K 1998. Comparison of intranasal
and intramuscular immunization against human immunodeficiency virus type I
with a DNA monophosphoryl lipid A adjuvant vaccine. Infect Immun 66:
823-826.
- Walker CM 1996. Cytotoxic T-lymphocyte responses
to the hepatitis C virus in humans and chimpanzees. Virology 7: 13-21.
- Wang S, Liu X, Fisher K, Smith JG, Chen F,
Tobery TW, Ulmer JB, Evans RK, Caulfield MJ 2000. Enhanced type I immune response
to hepatitis B DNA vaccine by formulation with calcium-or aluminum phosphate.
Vaccine 18: 1227-1235.
- Weiner GJ, Liu HM, Wooldridge JE, Dahle CE,
Krieg AM 1997. Immunostimulatory oligodeoxynucleotides containing the CpG
motif effective as immune adjuvants in tumor antigen immunization. Proc
Natl Acad Sci 94: 10833-10837.
© 2002
Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02017t1.jpg]
[oc02017f2.jpg]
[oc02017f1.jpg]
|

{kind=link}
{kind=link}
{kind=link}