
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 1, 2002, pp. 101-103
|
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(1) 2002, pp. 101-103
IgG Anti-IgE Autoantibodies
in Visceral Leishmaniasis
AM Atta/+, MLB Sousa-Atta, A D'Oliveira*,
RP Almeida*, MI Araújo*, EM Carvalho*
Departamento de Análises Clínicas
e Toxicológicas, Faculdade de Farmácia, Universidade Federal da
Bahia, Rua Barão de Geremoabo s/n°, Campus Ondina, 40171-970 Salvador,
BA, Brasil *Serviço de Imunologia, Hospital Universitário Prof.
Edgar Santos, Salvador, BA, Brasil
+Corresponding author. Fax: +55-71354.3862. E-mail: ajatta@ig.com.br
Received 5 March 2001
Accepted 3 October 2001
Code Number: oc02018
Procedures for IgG depletion in visceral leishmaniasis
(VL) and schistosomiasis sera using Sepharose-protein G beads also deplete IgE.
In this study, the presence of IgG anti-IgE autoantibodies in sera from patients
with VL (n = 10), and hepatic-intestinal schistosomiasis (n = 10) and from healthy
individuals (n = 10) was investigated. A sandwich ELISA using goat IgG anti-human
IgE to capture serum IgE and goat anti-human IgG peroxidase conjugate to demonstrate
the binding of IgG to the IgE captured was performed. VL sera had higher titers
(p < 0.05) of IgG anti-IgE autoantibodies (OD = 2.01 ± 0.43) than
sera from healthy individuals (OD = 1.35 ± 0.16) or persons infected
with Schistosoma mansoni
(OD = 1.34 ± 0.18). The immunoblotting carried out with eluates from
Sepharose-protein G beads used to deplete IgG from these sera and goat anti-human
IgE peroxidase conjugate, showed a similar pattern of bands, predominating the
75 kDa e-heavy
chain and also polypeptides resulting from physiological enzymatic digestion
of IgE. A frequent additional band immediately above 75 kDa was observed only
in VL sera.
Key words: IgE - IgG anti-IgE - protein G - visceral
leishmaniasis - schistosomiasis mansoni
Immunoglobulin E is an isotype of antibody that
has Fc receptors of different affinity on basophils, mast cells, eosinophils,
macrophages and platelets. The immune reaction of IgE fixed on these cells with
the corresponding antigen can release bioactive cell mediators as histamine,
sulfidopeptide leukotrienes, prostaglandin D2 or platelet-activating
factor, causing systemic or local inflammatory reactions. The involvement of
IgE in host immune response to parasite antigens was showed immediately after
its immunochemical identification in 1969, when high concentration of this protein
in serum was obtained from persons suffering of helminthiasis (Bennich et al.
1969, Kojima et al. 1972). The recent studies on the dichotomy of immune response
in Th1 and Th2 (Coffman et al. 1991), which is related to the pattern of cytokines
produced during antigenic stimulation, have shown that IgE is the mainly antibody
induced by IL-4 and IL-13 during a Th2 immunologic event (Romagnani 1998, 2000,
Hunter & Reiner 2000). This fact has provoked the interest in the research
of this isotype of immunoglobulin in several diseases associated to a Th2 immunological
pattern such as systemic lupus erythematosus and viral and parasitic infections
(Elkayam et al. 1995, Mazza et al. 1995, Atta et al. 1998). A common pitfall
in serum IgE immunoassays to diagnosis infections by protozoa or helminth, also
present in IgM tests, corresponds to false negative results originated by serologic
competition of IgG antibodies, generally present in greater concentration, and
affinity for common epitopes on target antigens. In order to solve this problem,
procedures of serum IgG depletion have been advised, employing anti-human IgG
antibodies obtained through immunization of different species, followed by elimination
of the immune precipitates formed after absorption by centrifugation. Staphylococcus
protein A, Streptococcus protein G or anti-IgG antibodies have also been
used as ligands in experiments of serum IgG depletion after conjugation with
inert particles as Sepharose beads.
Recently, when we investigated the occurrence
of IgE anti-Leishmania chagasi antibodies in sera from patients with
visceral leishmaniasis (VL), and also IgE anti-Schistosoma mansoni antibodies
in sera from individuals with schistosomiasis, we observed that the use of Sepharose-protein
G for the depletion of IgG caused significant fall in sensitivity of the specific
immunoassays when the results obtained were compared with those derived from
the use of RF-Absorbent (Behring Diagnostics, USA), a solution of purified anti-human
IgG antibodies (Sousa-Atta et al. 1999). Considering that IgG anti-IgE autoantibodies
have been demonstrated in healthy individuals and in patients with asthma, autoimmune
and parasite diseases (Inganas et al. 1981, Quinti et al. 1986, Gruber et al.
1988, Scheuer et al. 1991), and complexed IgG anti-IgE autoantibodies were documented
in sera from Crohn's disease patients and from patients with rheumatoid arthritis
(Huber et al. 1998, Millauer et al. 1999), the aims of this study was to evaluate
the levels of complexed IgG anti-IgE autoantibodies in sera from VL and from
with hepatic-intestinal schistosomiasis mansoni patients and also to investigate
if treatment of human serum with Sepharose-Protein G causes IgE depletion through
the sequestration of immunocomplexes IgG-IgE by the beads.
MATERIALS AND
METHODS
Sera - Sera from 10 VL and 10 schistosomiasis
mansoni patients were obtained from persons with clinical and laboratory diagnosis
of these parasitic diseases, all assisted by the staff of the Immunology Service
of the Hospital Universitário Professor Edgard Santos. Control sera were
from 10 healthy individuals having negative serology for rheumatic and infectious
diseases (American trypanosomiasis, viral hepatitis and syphilis).
Methods - The immunoassay to detect complexed
IgG anti-IgE autoantibodies was an ELISA of immunocapture of IgE, developed
on polystyrene microplates containing wells covered with goat IgG anti-IgE (Sigma
Chemical Co., USA) and a goat IgG anti-human IgG peroxidase conjugate, from
the same source, to demonstrate the im-munecomplexes IgG-IgE captured. Briefly,
the immune reaction to capture IgE was performed incubating 100 µl of
human sera diluted at 1/6 in 50 mM Tris-HCl (pH 7.5) buffered-saline containing
1% bovine serum albumine and 0.05% Tween 20 for 1 h at room temperature, while
the reaction with 100 µl of the diluted conjugate was developed in the
same conditions, after wash of the wells. The reactions were revealed with hydrogen
peroxide plus OPD (ortho-phenylenediamine) during 30 min, stopped with
2N HCl and determined at 492-600 nm in a DIAMEDIX BP-12 Microassay apparatus.
To demonstrate the sequestration of complexed
IgG anti-IgE by protein G, three sera from each group under study were diluted
at 1/5 in PBS containing 25% of Sepharose-protein G (Pharmacia Biotech, Uppsala,
Sweden) and incubated during 15 min at room temperature. After repeated wash
by centrifugation with PBS, the beads were treated with SDS-polyacrylamide gel
electrophoresis sample buffer containing 2-mercaptoethanol to elute the captured
material, followed by electrophoresis on a 10% acrylamide minigel. The polypeptides
fractionated were transferred by electrophoresis to a PVDF membrane (Immobilon,
Millipore, and USA) and analyzed for IgE by incubation with goat anti-human
IgE peroxidase conjugated (Sigma Chemical Co., USA). After new wash, the immune
reactions were revealed incubating the membrane with hydrogen peroxide plus
DAB (3,3´-diaminobenzidine), as usually.
Serum IgE concentration was determined by ELFA
using the Vidas 30 immunoanalyzer from biolabMeriéux.
Statistical analysis were performed through the
Primer PC statistic program using a non-parametric test U of Mann-Whitney.
RESULTS
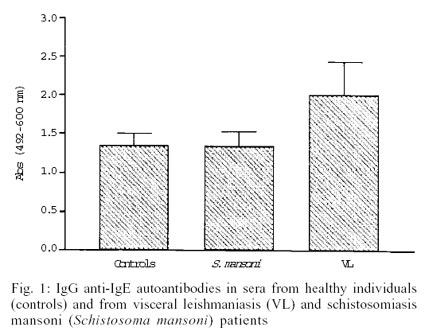
Complexed IgG anti-IgE autoantibodies were detected
in all sera studied. The results of the immunoassays to detect these immune
complexes are presented in the Fig. 1.
While schistosomiasis and normal sera presented similar titers, sera from VL
patients had higher levels of complexed IgG anti-IgE when compared to these
groups, as assessed by the U test of Mann-Whitney (p < 0.05). There was no
correlation between IgE concentration and complexed IgG anti-IgE level in serum
(p > 0.05). Patients from schistosomiasis group with serum IgE concentration
of 97 IU/ml or 5800 IU/ml presented similar levels of complexed IgG anti-IgE
autoantibodies.
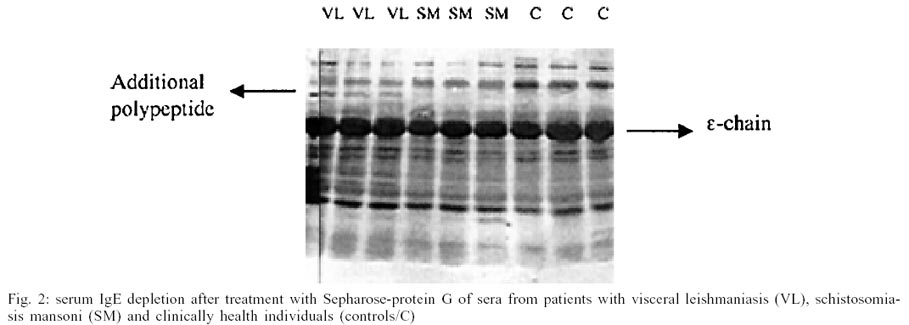
Immunochemical analysis carried out with material
eluted from batching experiments to deplete IgG from these sera evidenced significant
capture of IgE immunoglobulin during IgG binding to Sepharose protein G beads,
as demonstrated by intense brown colored band on PVDF membrane, with molecular
weight corresponding to 75 kDa IgE heavy chain in all sera analyzed, inclusive
from healthy controls. Other peptides of different weights that probably correspond
to the products of physiologic degradation of this immunoglobulin were also
observed (Fig. 2). However, only visceral
leishmaniasis sera presented an additional polypeptide with molecular weight
immediately above 75kDa-heavy chain.
DISCUSSION
The observations here reported indicated that
complexed IgG anti-IgE might be detected in high concentration in sera from
VL patients. These immune complexes are not increased in sera from patients
suffering from hepatic-intestinal schistosomiasis mansoni, and their titers
were not correlated with the elevated concentration of serum IgE presented by
some infected patients. On the other hand, sera from schistosomiasis group presented
the same reactivity pattern for IgG anti-IgE observed in sera from healthy controls,
which had IgE concentration below 150 IU/ml. This argues against the participation
of polyclonal B-cell activation in the production of IgG anti-IgE autoantibodies
and indicates that the biological significance of the increased levels of IgG
anti-IgE autoantibodies needs to be elucidated. According to previous hypothesis
IgG anti-IgE autoantibodies may exert the role of specific feed back molecules
that neutralize the IgE immune response induced by the cytokine network (Stadler
et al. 1993a). IgG anti-IgE autoantibodies are a heterogeneous population of
antibodies that execute functions of physiological control of the specific IgE
Th2 immune response, as inhibition of IgE synthesis and removal of IgE from
CD23 receptor (Stadler et al. 1993b). In atopic diseases as asthma these autoantibodies
have been implicated in the modulation of the IgE immune response, mainly because
they may react with epitopes located in C epsilon 2 domain of IgE, which is
involved in the binding of this immunoglobulin on Fce high affinity receptor
(FceRI) on the surface of basophils and mast cells, preventing the sensitization
of these cells (Shakib & Powell-Richards 1991, Stadler et al. 1995).
Immunoblotting analysis carried out with material
eluted from Sepharose-protein G beads used in serum IgG depletion confirmed
the presence of IgG anti-IgE autoantibodies in all sera tested and evidenced
significant cleavage of IgE under physiologic conditions. A frequent 75 kDa
Mr band corresponding to IgE e-heavy chain and polypeptides of different size,
all carrying e-heavy chain epitopes, were revealed with specific goat IgG anti-human
IgE peroxidase conjugate in the blots carried out with sera from the three groups,
including healthy control sera. Apparently, such pattern of IgE degradation
may reflect the action of intrinsic regulatory mechanisms used by the organism
to block the formation of circulating immune complexes with IgG anti-IgE autoantibodies,
avoiding therefore inflammation and autoimmunity. The observation of an unusual
fragment of IgE only in VL sera needs to be investigated. Current studies are
evaluating the participation of L. chagasi proteases in enzymatic digestion
of this immunoglobulin during infection, after binding to CD23 receptor on parasited
macrophages.
The information that IgG anti-IgE autoantibodies
contribute significantly for serum immunoglobulin E loss in laboratory procedures
of immunoglobulin G depletion using immobilized specific ligands is a relevant
subject in immunodiagnosis assays. The main consequence of this sequestration
of immune complexes formed by IgG and IgE, is an accentuated fall in sensitivity
of the immunoassays projected to detect specific serum IgE antibodies in vitro
and consequently occurrence of false negative results. Additionally, this finding
determine caution during the interpretation of IgG antibody positive tests obtained
with solid phase immunoassays covered with immobilized antigens and sera presenting
concomitant high concentrations of IgE specific antibodies and IgG anti-IgE
autoantibodies, as observed in Th2 immune responses from allergic or parasitic
diseases.
REFERENCES
- Atta AM, D'Oliveira Jr A, Correa J, Atta MLB,
Almeida RP, Carvalho EM 1998. Antileishmanial IgE antibodies: a marker of
active disease in visceral leishmaniasis. Am J Trop Med Hyg 59:
426-430.
- Bennich H, Ishizaka K, Ishizaka T, Johansson
SG 1969. A comparative antigenic study of gE-globulin
and myeloma-IgND. J Immunol 102: 826-831.
- Coffman RL, Varkila K, Scott P, Chatelain
R 1991. Role of cytokines in the differentiation of CD4+ T cells subsets in
vivo. Immunol Rev 123: 1-19.
- Elkayam O, Tamir R, Pick AI, Wysenbeek A 1995.
Serum IgE concentrations, disease activity and atopic disorders in systemic
lupus erythematosus. Allergy 50: 94-96.
- Gruber BL, Kaufman LD, Marchese MJ, Roth W,
Kaplan AP 1988. Anti-IgE autoantibodies in systemic lupus erythematosus. Arthritis
& Rheumatism 31: 1000-1006.
- Huber A, Genser D, Spitzauer S, Scheiner O,
Jensen-Jarolim E 1998. IgE/anti-IgE immune complexes in sera from patients
with Crohn's disease do not contain food-specific IgE. Int Arch Allergy
Immunol 115: 67-72.
- Hunter CA, Reiner SL 2000. Cytokines and T
cells in host defense. Cur Opin Immunol 12: 413-418.
- Inganas M, Johansson SGO, Bennich H 1981.
Anti-IgE autoantibodies in human serum: occurrence and specificity. Int
Arch Allergy Appl Immunol 65: 51-56.
- Kojima S, Yokagawa M, Tada T 1972. Raised
levels of serum IgE in human helminthiasis. Am J Trop Med Hyg 21: 913-918.
- Mazza DS, Grieco MH, Reddy MM, Meriney D 1995.
Serum IgE in patients with human immunodeficiency virus infection. Ann
Allergy Asthma Immunol 74: 411-414.
- Millauer N, Zuercher AW, Miescher SM, Gerber
HA, Seitz M, Stadler BM 1999. High IgE in rheumatoid arthritis (RA) patients
is complexed with anti-IgE autoantibodies. Clin Exp Immunol 115: 183-188.
- Quinti I, Brozek C, Wood N, Geha RS, Leung
DYM 1986. Circulating IgG autoantibodies to IgE in atopic syndromes. J
Allergy Clin Immunol 77: 586-594.
- Romagnani S 1998. T cell subsets (Th1, Th2)
and cytokines in autoimmunity. In NR Rose, IR Mackay (eds), The Autoimmune
Diseases, Academic Press, London, p. 163-191.
- Romagnani S 2000. T-cell subsets (Th1 versus
Th2). Ann Allergy Asthma Immunol 85: 9-18.
- Scheuer A, Haas H, Schlaak M 1991. Prevalence
and subclass distribution of IgG-anti-IgE autoantibodies in atopy and parasitosis.
Int Arch Allergy Appl Immunol 96: 271-276.
- Shakib F, Powell-Richards A 1991. Elucidation
of the epitopes locations of human autoanti-IgE: recognition of two epitopes
located within the C epsilon 2 and the epsilon 4 domains. Int Arch Allergy
Appl Immunol 95: 102-108.
- Sousa-Atta MLB, Araújo MI, D'Oliveira
Jr A, Ribeiro de Jesus A, Almeida RP, Atta AM, Carvalho EM 1999. Detection
of specific IgE antibodies in parasite diseases. Braz J Med Biol Res
32: 1101-1105.
- Stadler BM, Stampfli MR, Miescher S, Furukawa
K, Vogel M 1993a. Biological activities of anti-IgE antibodies. Int Arch
Allergy Immunol 102: 121-126.
- Stadler BM, Stampfli MR, Miescher S, Rudolf
M, Vogel M 1995. Cloning of human anti-IgE autoantibodies and their role in
regulation of IgE synthesis. Int Arch Allergy Immunol 107: 48-50.
- Stadler BM, Stampfli MR, Vogel M, Aebischer
I, Furukawa K, Holzner ME, Rudolf MP, Miescher S 1993b. A specific feedback
by anti-IgE autoantibodies on the cytokine network in allergy. Agents and
Actions 40 (Suppl.): 144-152.
© 2002
Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02018f1.jpg]
[oc02018f2.jpg]
|

{kind=link}
{kind=link}