
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 1, 2002, pp. 113-117
|
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(1) 2002, pp. 113-117
Detection and Characterization
of Rotavirus G and P Types from Children Participating in a Rotavirus Vaccine
Trial in Belém, Brazil
JDP Mascarenhas/+, AC Linhares, YB
Gabbay, JPG Leite*
Instituto Evandro Chagas, Fundação
Nacional de Saúde, 66090-000 Belém, PA, Brasil *Departamento de
Virologia, Instituto Oswaldo Cruz-Fiocruz, Rio de Janeiro, RJ, Brasil
+Corresponding author. Fax: +55-91-214.2005. E-mail: joanamascarenhas@iec.pa.gov.br
This work was supported by Instituto
Evandro Chagas-Funasa, Instituto Oswaldo Cruz-Fiocruz, Coordenação
Geral de Laboratórios-Funasa, and CNPq.
Received 15 March 2001
Accepted 3 October 2001
Code Number: oc02021
This study sought the characterization of
rotaviruses in a trial with a tetravalent rhesus-human rotavirus vaccine in
Belém, Brazil in children who received three doses of vaccine or placebo
in the 1st, 3rd and 5th months of life. Rotavirus electropherotypes, subgroups,
G serotypes, G, [P] and [P],G genotypes were determined in 93.3%, 95.9%, 93.3%,
73.3%, 95.5% and 92.2% of isolates, respectively. Serotypes G1, G2 and G4 were
detected in 58.9%, 30% and 4.4% of the cases, respectively. Rotavirus genotype
G5 was detected for the first time in Northern region in 4.4% of the infections.
Rotavirus genotypes P[8], P[4], P[6] and P[8+6] were detected in 54.5%, 26.7%,
12.2%, and 2.2% of the cases, respectively. The predominant genotypes were P[8],G1
and P[4],G2 with 53% and 26.6% of the infections, respectively. Unusual strains
accounted for 20.5% including P[4],G1, P[6],G1, P[6],G4, P[6],G5, P[8],G2, P[8],G5.
Mixed infections involving P[8+6],G2 and P[8+6],G1 were also noted. The neonatal
P[6] strains associated with diarrhea were detected among children aged 9-24
months. To our knowledge, this study represents the first in Brazil to analyse,
on molecular basis, rotavirus genotypes from children participating in a rotavirus
vaccine trial. These results are of potential importance regarding future rotavirus
vaccination strategies in Brazil.
Key words: rotavirus - vaccine - genotype G -
genotype P - Belém - Brazil
Rotaviruses A constitute the most important cause
of severe gastroenteritis among infants and young children in developing and
developed countries, accounting for about 680,000 deaths per year in developing
countries (Kapikian & Chanock 1996, Miller & McCann 2000). Because of
the high mortality-rate associated with rotavirus diarrhoea, particularly in
the developing countries, the availability of an effective vaccine is a goal
to be pursued.
The rotavirus belongs the Reoviridae family,
genus Rotavirus. The complete viral particle is constituted by a triple-layered
shell protein and the genome that consists of 11 segments of double-stranded
RNA (dsRNA) each one coding a protein (Kapikian & Chanock 1996). The proteins
VP4 and VP7 induce type-specific neutralising antibodies and are involved with
the immunity protection. G serotype is associated with VP7 protein whereas P
serotype refers to VP4 protein (Kapikian & Chanock 1996).
Fourteen G serotypes/genotypes, of which 10 (G1-G6,
G8-G10 and G12) were described infecting humans (Parashar et al. 1998), and
20 P genotypes, as specified by the 4th dsRNA segment; 4 of these were described
infecting humans, as follows: P[4], P[6], P[8], and P[9] (Estes 1996). The serotypes
G1-G4, are the most common cause of disease worldwide (Kapikian & Chanock
1996, Gentsch et al. 1996). In developing countries other G serotypes/genotypes
are important such as G5, G6, G8, G9, and G10 (Gentsch et al. 1996, Santos et
al. 1998).
On the basis of the current binary proposed system
for rotavirus characterization, the majority of isolates from diarrhoeic children
fall into four groups: P[8],G1, P[4],G2, P[8],G3, and P[8],G4 (Bishop et al.
1991, Woods et al. 1992, Beards et al. 1995). Of these, P[8],G1 and P[4],G2
rotavirus strains are the most prevalent worldwide (Rasool et al. 1993, Das
et al. 1994, Santos et al. 1994, Gentsch et al. 1996).
The present report documents the diversity of
rotavirus G and P types from children participating in trial using the tetravalent
rhesus-human reassortant rotavirus vaccine (RRV-TV) in Belém, Brazil.
MATERIALS
AND METHODS
Patients and methods - This study was
aproved by the Regional Council of Medicine of the State of Pará, the
Secretary of Public Health of the State of Pará, and the Ministry of
Health of Brazil, and by the Ethical Commitee of Instituto Evandro Chagas (Belém)
and Ethical Review Commitee of the World Health Organization.
The study was a 2-year, prospective, double-blind,
placebo-controlled, randomized trial in which infants received three doses of
RRV-TV or placebo in the 1st, 3rd and 5th months of life (Linhares
et al. 1996). In the present study we are considering 90 rotavirus positive
cases diagnosed by enzyme immunoassay (DAKOTM, Copenhagen, Denmark),
involving 83 children with gastroenteritis wich received three doses of RRV-TV
vaccine (270 children) or placebo (270 children).
G serotyping and subgrouping by enzyme immunoassay
- Subgrouping and G-serotyping of rotavirus-positive samples were performed
essentially as described by Taniguchi et al. (1987), by using monoclonal antibodies
against each of the subgroups I and II and human G1, G2, G3, and G4 serotypes,
which were kindly provided by Dr Shozo Urasawa, Department of Hygiene and Epidemiology,
Sapporo Medical College, Sapporo, Japan.
Polyacrylamide gel electrophoresis - The
double stranded RNA was extracted from 400 µl of positive suspensions
follow the metodology described by Boom et al. (1990). The electrophoresis was
performed using a 5% polyacrylamide slab gels (PAGE) and the gels were then
stained with silver nitrate as previously described by Pereira et al. (1983).
Reverse transcription-polymerase chain reaction
for G and P rotavirus genotyping - The reverse-transcription followed by
polymerase chain reaction (RT-PCR) for G and P rotavirus genotyping was performed
in two steps, as described by Gouvea et al. (1990, 1994) and Gentsch et al.
(1992), respectively, with modifications introduced by Leite et al. (1996).
In the first amplification, a mixture of consensual primers 9con1/9con2 or Beg/End9
(G genotype) and 4con2/4con3 (P genotype) was used. To assess both G and P broad
reactivity the mixture was cycled in a termocycler (Perkin Elmer, GeneAmp PCR
System 9600), being submitted to 30 cycles of amplification. The amplicons were
then eletrophoresed on 1% agarose gel in Tris-Borate-EDTA buffer (TBE) in Wide
Mini-Sub Cell GT (Bio-Rad Laboratories, Hercules, CA, USA). The gel was stained
with ethidium bromide (0.5 µg/ml) and cDNAs visualized and photographed
using a system Gel Doc 1000 (Bio-Rad, Laboratories, Hercules, CA, USA).
The nested-PCR was carried out using a mixture
of G (G1-G5 and G9 and rotavirus animal primers Beg 9, d end 9, CRW8, end 9-6)
(Gouvea et al. 1990, 1994), and P (P[4], P[6], P[8], and P[9]) (Gentsch et al.
1992) specific genotyping oligonucleotide primers by 30 cycles of amplification.
All amplicons were subsequently subjected to electrophoresis on 1% agarose in
TBE buffer containing ethidium bromide (0.5 µg/ml).
Southern hybridization and chemiluminescent
detection - Southern hybridization with oligonucleotide probes was carried
out to confirm the P (Ramachandran et al. 1996) and G (Leite et al. 1996) genotype-specificities
according to the method described by Ando et al. (1995), with modifications
introduced by Leite et al. (1996). Dig-probes were used to discriminate between
porcine G5 OSU-like strains from G5 human strains isolated from Brazilian infants,
as described by Alfieri et al. (1996).
RESULTS
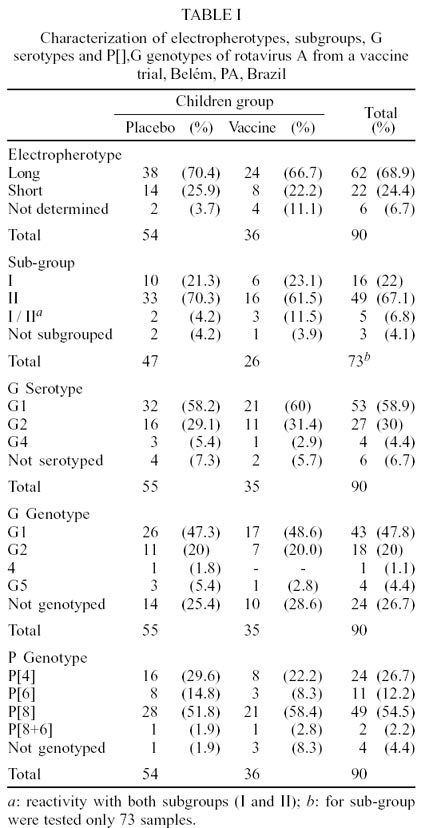
The PAGE showed a clear profile in 84 (93.3%)
out of 90 positive samples. The predominant electropherotype was long, accounting
for 38 (70.4%) and 24 (66.7%) of tested samples in the placebo and vaccine groups
(Table I).
The sub-group specificity was carried out in
70 out of 73 (95.9%) positive samples, of which, sub-group II was the most prevalent,
followed by sub-group I in both groups (Table
I).
Serotyping was carried out on 84 (93.3%) out
of 90 rotavirus positive specimens. G1, G2 and G4 were detected and the most
prevalent serotype was G1 in both placebo (58.2%) and vaccine (60%) groups (Table
I). The usual correlation between subgroups and serotypes was noted in 80%
of positive samples.
Nested-PCR was carried out in 90 samples to characterize
the G rotavirus genotypes. Using 9con1/9con2 specific primers it was possible
to characterize rotavirus G genotype in 66 (73.3%) of isolates. From 66 samples
with defined G genotype, 43 (47.8%) were genotype G1, 18 (20%) G2, 1 (1.1%)
G4 and 4 (4.4%) G5.
From the 24 remaining samples without G genotype,
19 with available specimens were tested using rotavirus animal primers Beg 9,
d end 9, CRW8, end 9-6, obtained from rotavirus of animal origin, being all
negatives.
From 86 (95.5%) samples with defined P genotypes,
24 (26.7%) were classified as genotype P[4], 11 (12.2%) P[6], and 49 (54.5%)
P[8]. Two samples (2.2%) showed mixed infections P[8+6] (Table
I). The P[6] genotype was observed in both placebo and vaccine groups involving
diarrhoeic children with ages between 9 and 24 months (data not shown).
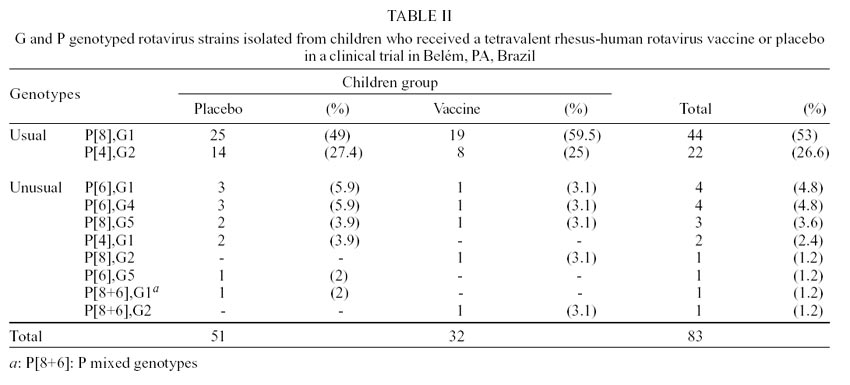
The binary characterization [P],G was possible
in 83 (92.2%) out of 90 positive samples. The predominant combination was the
usual genotypes: P[8],G1 (53%) and P[4],G2 (26.6%). Unusual genotypes or mixed
infections were observed in 20.5% of strains, such as: P[6],G1 (4.8%), P[6],G4
(4.8%), P[8],G5 (3.6%), P[4],G1 (2.4%), P[8],G2 (1.2%), P[6],G5 (1.2%), P[8+6],G1
(1.2%) and P[8+6],G2 (1.2%) (Table II).
Southern-hybridization using specific G1, G2,
and G5 probes was carried out in 21 amplicons. Eight amplicons with expected
size of genotype G1 and four with expected size of genotype G2 were confirmed
as genotype G1 and G2 with the specific probes. However, only 4 out of 9 amplicons
with expected size for G5 (742bp) were positive by Southern-hybridization (data
not shown). Similarly, 11 samples were characterized by Southern-hybridization
to P genotypes and, four reacted specifically with the probe P[6], three with
probe P[8], two with probe P[4] and two with both probes P[8] and [6] (data
not shown).
DISCUSSION
This is the first study in Brazil to determine
the G and P rotavirus genotypes circulating among children participating in
a vaccine trial carried out in Belém. To our knowledge, characterization
of P genotypes has not been performed in previous vaccine trials worldwide (Clark
et al. 1995, Lanata et al. 1996, Linhares et al. 1999).
In the present investigation it was observed
an usual correlation of subgroup/serotype in 80% of positive cases. Similar
rates were reported by Mohamed et al. (1994) in a study involving infants and
young children with acute gastroenteritis in Saudi Arabia.
In this study, a high proportion (93.3%) of samples
were G-serotyped using monoclonal antibodies. This rate is higher than those
observed in previous studies carried out in Belém and Goiânia,
Brazil, where only 50% and 61% of samples could be serotyped (Linhares et al.
1988, Cardoso et al. 2000). It is likely that examination shortly after collection
has accounted for the higher rates of G-serotyping as compared to G- genotyping.
This seems to be an unusual result since almost all published studies (including
those from the Brazilian groups) found that RT-PCR typing greatly improves rotavirus
G typing. Using primers Beg/End9 to amplify animal rotavirus strains in remaining
19 samples unsuccessfully amplified, we could postulated that those samples
had mismatches in the consensual region for primer hybridization.
Among the four more epidemiologically important
rotavirus G serotypes worldwide, three were identified in the present study:
G1, G2 and G4. In addition, rotavirus genotype G5, not detected previously in
the Northern Region of Brazil, occurred in 4.4% of isolates.
Santos et al. (1998), studying 49 rotavirus positive
samples from diarrhoeic children from Rio de Janeiro, Brazil, found usual G1
and G3 genotypes in 27% and 12% of infection cases, followed by 61% of uncommon
genotypes as G5 (25%), G10 (16%), G8 (5%), and mixed G types (16%). In the present
investigation, the serotype G1 was detected in 58.9% of tested strains, whereas
no G3 was recorded. In contrast, rotavirus serotypes G2 and G4 accounted for
30% and 4.4% of isolates, respectively. The results obtained in the present
study were similarly to those obtained in the United Kingdon by Beards and Graham
(1995), who identified G1, G2, G3, and G4 in 93% of isolates.
Linhares et al. (1988), in a study carried out
in Belém from December 1982 to March 1986, showed that serotype G1 was
the most prevalent (50%), followed by G2 (30%), G4 (17%) and G3 (3%). In addition,
Cardoso et al. (2000), during study conducted in Goiânia, from 1987 to
1994 recorded the following rotavirus serotypes: G1 (32%), G2 (46%), G3 (16%),
G4 (2%), and G5 (4%).
This investigation showed relative frequencies
of genotypes which differ from those found in another study carried out in Belém,
from November 1992 to November 1994, when rotavirus serotype G2 was largely
prevalent (80%) over the other serotypes in nosocomial infections (Gusmão
et al. 1995).
The rotavirus genotype G5 was identified in 1991
(two isolates) and 1992 (two isolates), being detected at rates of 4.4%, three
in the placebo and one in the vaccine group. Three samples were characterized
as genotype P[8],G5 and one as genotype P[6],G5. The five remaining samples
with expected size for G5, were negative by hybridization. Probably those samples
had mismatches in the consensual region at probe hybridization since no amplification
could be yielded.
To date, serotype G5 has largely been detected
in pigs and, at lower rates, in equines, and also in humans. Gouvea et al. (1994)
detected serotype G5 in 38 faeces of the 329 samples (12%) for Brazilian children
with diarrhoea. Leite et al. (1996) recorded the genotype G5 circulating in
several states of Brazil, suggesting a broad distribution of this unusual genotype
in Brazil. Previous multicentric studies in Brazil have shown that rotavirus
bearing P[8],G5 type-specificity may account for up to 9% of cases of acute
gastroenteritis among infants and young children (Timenetsky et al. 1994, Leite
et al. 1996, Cardoso et al. 2000).
Alfieri et al. (1996) by Southern-hybridization,
sequencing, and RNA-RNA hybridization analysis, showed that rotavirus genotype
G5 circulating in Brazil has homology with human rotavirus genotype P8 (Wa-like)
and porcine rotavirus genotype G5 (OSU-like). These results suggest that these
strains have naturally reassorted, involving members of both P8 (Wa) and G5
(OSU) rotavirus genogroups.
With regards to the characterization of P genotypes
in the present study, P[8], P[4] and P[6] accounted for 54.5%, 26.7% and 12.2%
of isolates, respectively. These results were similar to those observed in South
Africa, where genotypes P[8], P[4] and P[6] occurred in 64%, 22% and 8% of cases,
respectively (Mphahlele & Steele 1995).
Working with hospitalized children in Belém,
Mascarenhas et al. (1999) characterized 86% of isolates, in contrast with the
present investigation when genotype P characterization was obtained in 95.5%
of isolates.
Neonatal P[6] strains had been previously identified
among asymptomatic neonates and has been regarded as avirulent (Bishop et al.
1983, Haffejee 1991). The results of the present study are, however, in contrast
with those observations regarding two aspects: (i) P[6] was associated with
diarrhoeic cases; and (ii) involved children older than 28 days.
In this study, genotypes P[8],G1 and P[4],G2
occurred in 53% and 26.6% of isolates, respectively. Leite et al. (1996), studying
isolates from 9 states and the Federal District of Brazil, described the genotypes
P[8],G1, P[4],G2, P[8],G3 and P[8],G4, in 43%, 12%, 6% and 6% of isolates, respectively.
Ramachandran et al. (1998), in USA, detected P[8],G1, P[4],G2, and P[8],G3,
at rates of 66.4%, 8.3%, and 6.9% of samples, respectively.
In the present investigation it was possible
to characterize 79.5% and 20.5% of the samples as usual and unusual genotypes,
respectively. Leite et al. (1996), in a countrywide study in Brazil, found genotypes
P[8],G5, P[6],G2, P[9],G2 and P[9],G3 in 12% of the situations, and mixed infections
in 21% of cases. Ramachandran et al. (1998), analyzing samples in a multicenter
investigation in USA identified genotypes P[6],G9, P[8],G9, P[6],G1, P[8],G2
and P[4],G1 in 9.2% of the cases. In India Ramachandran et al. (1996) observed
unusual strains in 43%, of tested strains with results similar to our study,
if genotypes P[6],G1 and P[6],G4 are considered.
The rotavirus genotyping in Brazil demonstrated,
in general, a high rate of mixed infection and samples frequently untypeable.
These results shown a complexity of serotypes/genotypes G in children participating
in a trial with a candidate rotavirus vaccine.
The rotavirus vaccine (RRV-TV Rotashieldâ)
produced by Wyeth-Ayerst Research and licensed for use in the United States,
has been suspended recently, following the occurrence of intussusception among
vaccinated children (CDC 1999). A newly developed candidate rotavirus vaccine,
P[8],G1, of human origin, has been recently administered to children, resulting
in a rate of efficacy of about 90% against rotavirus gastroenteritis (Bernstein
et al. 1999). Large-scale field trials are currently under way in developing
countries.
These results described above have importance
regarding a future rotavirus immunization strategies in Brazil,
and underscore the need for a countrywide monitoring a rotavirus G- and P-types
before the introduction of a rotavirus vaccine.
ACKNOWLEDGEMENTS
To Dr Jon Gentsch and Dr Roger Glass, members
of the WHO/PAHO Rotavirus Collaborating Center in the Viral Gastroenteritis
Section of the Centers for Disease Control and Prevention for providing specific
primers and Drs Ronaldo Barros de Freitas, Consuelo Silva de Oliveira, Talita
Furtado Monteiro for clinical support in the follow-up of children and Mr Antônio
Miranda for technical support.
REFERENCES
- Alfieri AA, Leite JPG, Nakagomi O, Kaga E,
Woods PA, Glass RI, Gentsch JR 1996. Characterization of human rotavirus genotype
P[8]G5 from Brazil by probe-hybridization and sequence. Arch Virol
141: 2353-2364.
- Ando T, Monroe SS, Gentsch JR, Jin Q, Lewis
DC, Glass RI 1995. Detection and differentiation of antigenically distinct
small round-structured viruses (Norwalk-like viruses) by reverse transcription-PCR
and Southern hybridization. J Clin Microbiol 33: 64-71.
- Beards G, Graham C 1995. Temporal distribution
of rotavirus G-serotypes in the West Midlands region of the United Kingdom,
1983-1994. J Diarrhoeal Dis Res 13: 235-237.
- Beards GM, Desselberger U, Flewett TH 1995.
Temporal and geographical distribution of human rotavirus serotypes, 1983
to 1988. J Clin Microbiol 27: 2827-2833.
- Bernstein DI, Sack DA, Rothstein E, Reisinger
K, Smith VE, O'Sullivan D, Spriggs DR, Ward RL 1999. Efficacy of live, attenuated,
human rotavirus vaccine 89-12 in infants: a randomised placebo-controlled
trial. Lancet 354: 287-290.
- Bishop RF, Barnes GL, Cipriani E, Lund JS
1983. Clinical immunity after neonatal rotavirus infection: a prospective
longitudinal study in young children. N Engl J Med 309: 72-76.
- Bishop RF, Unicomb LE, Barnes GL 1991. Epidemiology
of rotavirus serotypes in Melbourne, Australia, from 1973 to 1989. J Clin
Microbiol 29: 862-868.
- Boom R, Sol CJA, Salimans MMM, Jansen CL,
Wertheim-Van Dillen PME, Van Der Noordaa J 1990. Rapid and simple method for
purifications of nucleic acids. J Clin Microbiol 28: 495-503.
- Cardoso DD, Soares CM, Azevedo MS, Leite JP,
Munford V, Racz ML 2000. Serotypes and subgroups of rotavirus isolated from
children in central Brazil. J Health Popul Nutr 18: 39-43.
- CDC-Centers for Disease Control and Prevention
1999. Withdrawal of rotavirus vaccine recommendation. MMWR 282: 1007.
- Clark HF, White CJ, Offit PA, Stiuson D, Eiden
J, Weaver S, Cho I, Shaw A, Krah D, Ellis R 1995. Preliminary evaluation of
safety and efficacy of quadrivalent human-bovine reassortant rotavirus vaccine.
Ped Res 37: 127A.
- Das BK, Gentsch JR, Cicirello HG, Woods PA,
Gupta A, Ramachandran M, Kumar R, Bhan MK, Glass RI 1994. Characterization
of rotavirus strains from newborns in New Delhi, India. J Clin Microbiol
34: 1820-1822.
- Estes MK 1996. Advances in molecular biology:
impact on rotavirus vaccine development. J Infect Dis 174 (Suppl.):
S37-S46.
- Gentsch JR, Glass RI, Woods P, Gouvea V, Gorziglia
M, Flores J, Das BK, Bhan MK 1992. Identification of group A rotavirus gene
4 types by polymerase chain reaction. J Clin Microbiol 30: 1365-1373.
- Gentsch JR, Woods PA, Ramachandran M, Das
BK, Leite JP, Alfieri A, Kumar R, Bhan MK, Glass RI 1996. Review of
G and P typing results from a global collection of rotavirus strains: implications
for vaccine development. J Infect Dis 174 (Suppl.): 30-36.
- Gouvea V, De Castro L, Do Carmo Timenetsck
M, Greenberg H, Santos N 1994. Rotavirus serotype G5 associated with diarrhea
in Brazilian children. J Clin Microbiol 32: 1408-1409.
- Gouvea V, Glass RI, Woods P, Taniguchi K,
Clark FH, Forrester B, Fang ZY 1990. Polymerase chain reaction amplification
and typing of rotavirus nucleic acid from stool specimens. J Clin Microbiol
28: 276-282.
- Gusmão RHP, Mascarenhas JDP, Gabbay
YB, Lins-Lainson Z, Ramos FLP, Monteiro TAF, Valente AS, Linhares AC 1995.
Rotaviruses as a cause of nosocomial, infantile diarrhoea in Northern Brazil.
Mem Inst Oswaldo Cruz 90: 743-749.
- Haffejee I 1991. Neonatal rotavirus infections.
Rev Infect Dis 13: 957-962.
- Kapikian AZ, Chanock RM 1996. Rotaviruses.
In BN Fields, DM Knipe, PM Howley (eds), Fields Virology, Lippincott-Raven,
Philadelphia, p.1657-1708.
- Lanata CF, Black RE, Flores J, Lazo F, Butron
B, Linares A, Huapaya A, Ventura G, Gil A, Kapikian AZ 1996. Immunogenicity,
safety and protective efficacy of one dose of the rhesus rotavirus vaccine
and serotype 1 and 2 human-rhesus rotavirus reassortants in children from
Lima, Peru. Vaccine 14: 237-243.
- Leite JPG, Alfieri AA, Woods PA, Glass RI,
Gentsch JR 1996. Rotavirus G and P types circulating in Brazil: characterization
by RT-PCR, probe hybridization, and sequence analysis. Arch Virol 141:
2365-2374.
- Linhares AC, Gabbay YB, Mascarenhas JDP, Freitas
RB, Flewett TH, Beards GM 1988. Epidemiology of rotavirus subgroups and serotypes
in Belém, Brazil: a three-year study. Ann Virol (Inst Pasteur) 139:
89-99.
- Linhares AC, Gabbay YB, Mascarenhas JDP, De
Freitas RB, Oliveira CS, Bellesi N, Monteiro TAF, Lins-Lainson Z, Ramos FLP,
Valente SA 1996. Immunogenicity, safety and efficacy of rhesus-human, reassortant
rotavirus vaccine in Belém, Brazil. Bull WHO 74: 491-500.
- Linhares AC, Lanata, CF, Hausdorff WP, Gabbay
YB, Black RE 1999. Reappraisal of the Peruvian and Brasilian lower titer tetravalent
rhesus-human reassortant rotavirus vaccine efficacy trials: analysis by severity
of diarrhea. Pediatr Infect Dis J 18: 1001-1006.
- Mascarenhas JDP, Gusmão RHP, Barardi
CRM, Paiva FL, Simões CO, Gabbay YB, Monteiro TAF, Linhares AC 1999.
Characterization of rotavirus P genotypes circulating among paediatric inpatients
in northern Brazil. Rev Inst Med Trop São Paulo 41: 165-170.
- Miller MA, McCann L 2000. Policy analysis
of the use of hepatitis B, Haemophilus influenzae type B, Streptococcus
pneumoniae-conjugate and rotavirus vaccines in national immunization schedules.
Health Econ 9: 19-35.
- Mohammed KA, El Assouli SM, Banjar ZM 1994.
Human rotavirus subgroups and serotypes in children with acute gastroenteritis
in Saudi Arabia from 1988 to 1992. J Med Virol 44: 237-242.
- Mphahlele MJ, Steele AD 1995. Relative frequency
of human rotavirus VP4 (P) genotypes recovered over a ten-year period from
South African children with diarrhoea. J Med Virol 47: 1-5.
- Parashar UD, Bresee JS, Gentsch JR, Glass
RI 1998. Rotavirus. Emerg Infect Dis 4: 561-570.
- Pereira HG, Azeredo RS, Leite JP, Barth OM,
Sutmoller F, De Farias V, Vidal MN 1983. Comparison of polyacrylamide gel
electrophoresis (PAGE), immuno-electron microscopy (IEM) and enzyme immunoassay
(EIA) for the rapid diagnosis of rotavirus infection in children. Mem Inst
Oswaldo Cruz 78: 483-490.
- Ramachandran M, Das BK, Vij A, Kumar R, Bhambal
SS, Kesari N, Rawat H, Bahl L, Thakur S, Woods PA, Glass RI, Bhan MK, Gentsch
JR 1996. Unusual diversity of human rotavirus G and P genotypes in India.
J Clin Microbiol 34: 436-439.
- Ramachandran M, Gentsch JR, Parashar UD, Jin
S, Woods PA, Holmes JL, Kirkwood CD, Bishop RF, Greenberg HB, Urasawa S, Gerna
G, Coulson BS, Taniguchi K, Bresee JS, Glass RI 1998. Detection and characterization
of novel rotavirus strains in the United States. J Clin Microbiol 36:
3223-3229.
- Rasool NBG, Larralde G, Gorziglia MI 1993.
Determination of human rotavirus VP4 using serotype-specific cDNA probes.
Arch Virol 133: 275-282.
- Santos N, Lima RCC, Pereira CFA, Gouvea V
1998. Detection of rotavirus types G8 and G10 among brazilian children with
diarrhea. J Clin Microbiol 36: 2727-2729.
- Santos N, Riepenhoff-Talty M, Clark HF, Offit
P, Gouvea V 1994. VP4 genotyping of human rotavirus in the United States.
J Clin Microbiol 32: 205-208.
- Taniguchi K, Urasawa T, Morita Y, Greenberg
HB, Urasawa S 1987. Direct serotyping of human rotavirus in stools by an enzyme-linked
immunosorbent assay using serotype 1-, 2-, 3-, and 4- specific monoclonal
antibodies to VP7. J Infect Dis 155: 1159-1166.
- Timenetsky MC, Santos N, Gouvea V 1994. Survey
of rotavirus G and P types associated with human gastroenteritis in São
Paulo, Brazil. J Clin Microbiol 32: 2622-2624.
- Woods PA, Gentsch JR, Gouvea V, Mata L, Santosham
M, Bai ZS, Urasawa S, Glass RI 1992. Distribution of serotypes of human
rotavirus in different populations. J Clin Microbiol 30: 781-785.
© 2002
Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02021t2.jpg]
[oc02021t1.jpg]
|

{kind=link}
{kind=link}