Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(1) 2002, pp. 127-132
Cercarial Chaetotaxy and Sex
Differentiation of Schistosoma mansoni Deriving from Humans and Nectomys
squamipes (Muridae: Sigmondontinae) in Brazil
Nilcéa Freire, José Roberto Machado-Silva/+,
Rosângela Rodrigues-Silva*, Luís Rey**/++
Departamento de Patologia e Laboratórios,
Faculdade de Ciências Médicas, Universidade do Estado do Rio de
Janeiro, Av. 28 de Setembro 87 fundos, 5º andar, 20551-001 Rio de Janeiro,
RJ, Brasil *Laboratório de Helmintos Parasitos de Vertebrados, Departamento
de Helmintologia **Laboratório de Biologia e Controle da Esquistossomíase,
Departamento de Medicina Tropical, Instituto Oswaldo Cruz-Fiocruz, Rio de Janeiro,
RJ, Brasil
+Corresponding author and CNPq and Faperj fellow. Fax: + 55-21-2587.6112.
E-mail: machado @uerj.br
++CNPq fellow
Received 3 May 2001
Accepted 28 August 2001
Code Number: oc02024
A comparative study was made between sympatric
isolates of Schistosoma mansoni:
one from a wild rodent (R) Nectomys
squamipes and another one from humans
(H) isolated from a low endemic schistosomiasis transmission area in Brazil.
Our purpose was to detect differences between them concerning chaetotaxy (number
and pattern of distribution of the argentophilic papillae) of the cercariae
by means of silver impregnation. No significant difference (x > 0.05) between
isolates was noted. Nevertheless, a significant difference (x < 0.05) was
observed in the cercarial index (ratio of the distance between the first and
the second preacetabular papillae and the distance between the first and the
second dorsal preacetabular papillae) of male and female cercariae in both isolates.
Males presented a greater cercarial index than females. By means of multivariate
analysis, male cercariae were distinguished from female cercariae through the
following characteristics: average number of dorsal papillae on the right quadrant,
average number of ventral middle papillae on the right quadrant (H isolate)
and average number of dorsal middle papillae on the left quadrant (R isolate).
The results suggest that R and H isolates belong to the same population that
could complete its life cycle in rodent-snail-rodent fashion.
Key words: Schistosoma mansoni - cercariae
chaetotaxy - sex determination - Nectomys squamipes - human host - schistosomiasis
mansoni - Brazil
In some areas of schistosomiasis mansoni transmission
in Africa and in the Antilles, certain rodent (R) species behave as hosts for
Schistosoma mansoni (Combes et al. 1976, Rodrigues e Silva et al. 1992).
In Guadeloupe three types of foci were identified: transmission exclusively
between humans (H), between H and R (amphixenosis) and exclusively between R
(Cassone et al. 1981). In Brazil some R species with semi-aquatic habits, with
different degrees of sinantropic, are considered as possible reservoirs for
S. mansoni (Rey 1993). To date, no transmission area exclusive to R has
been identified, however, the infection level of these hosts increases with
proximity to H housing (D'Andrea et al. 2000). Nonetheless, little is known
about the phenotypic characteristics of the larvae forms (cercariae) of the
strains of parasites isolated from these hosts.
Chaetotaxic study has allowed the characterization
of interspecific variations of S. mansoni. African strains present lateral
papillae in a triangular shape and a cercarial index of 1.0. They are thus different
from American strains where the disposition is linear and the index is greater
(1.0-1.4) (Bayssade-Dufour 1979b). Between two strains (JR e C5) from Venezuela,
it has been demonstrated that the former demonstrated a higher variation coefficient
in the total number of papillae (dorsal and ventral) and more than one anterior
ventral papilla (Pino et al. 1997). Variations have also been detected between
the cercarial indices of H (1.0-1.4) and murine (1.34-2.0) strains, in the S.
mansoni foci in Guadeloupe (Bayssade-Dufour 1977, Cassone et al. 1981).
Among the strains that exist in Brazil, it is
known that LE, SJ and another one from Recife (kept for several generations
under laboratory conditions) present significant differences in the cercarial
index (Cassone et al. 1978, Araújo et al. 1987). However, there are no
comparative chaetotaxic studies of cercariae from H and R isolates.
This article adds to a comparasion between the
morphological characteristics (Machado-Silva et al. 1994, Neves et al. 1998)
of sympatric isolates, derived from a low level endemic area (Rey 1993), where
schistosomiasis infection is present in the H population and in the wild R Nectomys
squamipes (Rodrigues e Silva et al. 1992, D'Andrea et al. 2000).
MATERIALS
AND METHODS
Isolates studied - The sympatric parasites
studied in this trial were obtained from the Pamparrão Valley, a rural
area in the municipality of Sumidouro (22°02'46''S; 42°41'21''W),
about 200 km from Rio de Janeiro City, State of the Rio de Janeiro, Brazil.
The isolates were studied in their first passage under laboratory conditions.
The H isolate was obtained from the stools of
autochthonous subjects and the R isolate from liver, stools or bowel contents
of 13 specimens of N. squamipes (2n = 56) (Bonvicino et al. 1996). The
isolation methodology and the maintenance in the laboratory have been presented
in another publication (Machado-Silva et al. 1994). Specimens from a sympatric
Biomphalaria glabrata colony, kept under laboratory conditions, were
exposed to unimiracidial infection of R or H isolates. The snails were kept
in glass aquariums and fed with fresh lettuce (Lacttuca sativa).
The cercariae eliminated after exposure of snails
to a light source, were stained with silver nitrate, placed on a glass slide
(Combes et al. 1976) and drawn in a light chamber (Olympus model BH2-DA). A
lens with amplification 40, a WHK 10X ocular and a NFK 5xLD photographic ocular
was used. A lens with 100 magnification with the same ocular and photographic
ocular was used in order to determine the cercarial index.
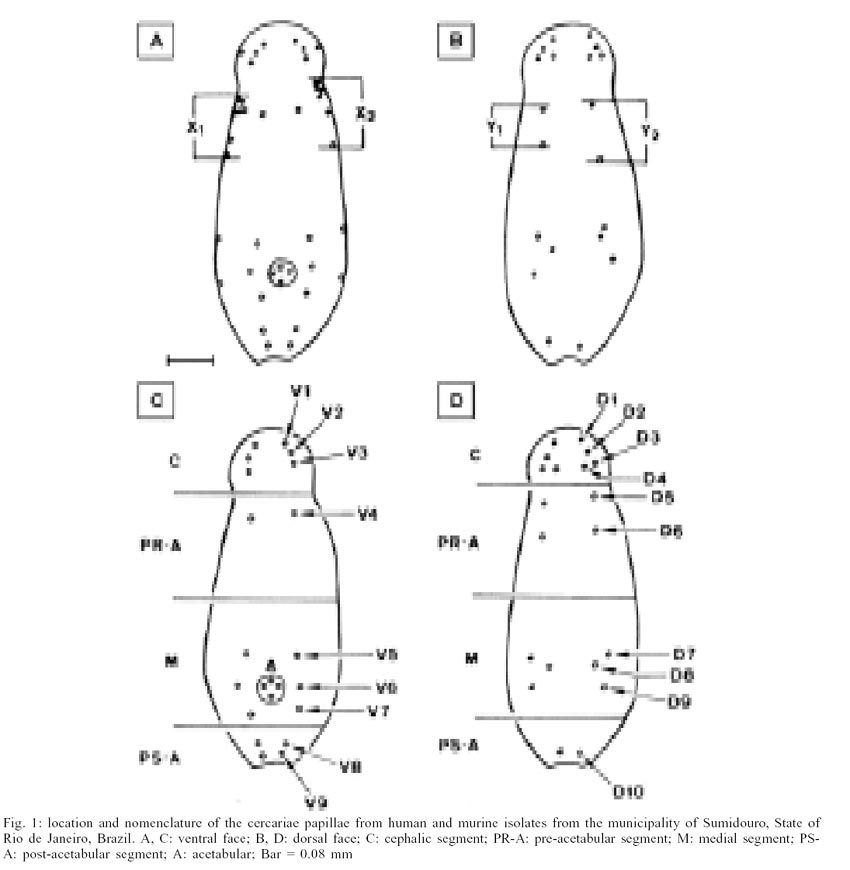
Calculation of the cercarial index - Defined
as the ratio of the distance between the first and the second preacetabular
papillae and the distance between the first and the second dorsal preacetabular
papillae (Fig. 1A, B) according to Bayssade-Dufour
(1979a): n = ½ ( x1/ y1 + x2/ y2
), were x1 and x2: distances between the first AIL papilla
and the AIIL papilla, for each of the cercarial hemi-bodies; and y1
and y2: distance between the AID papilla and AIID papilla, of each
cercarial hemi-body.
A lens with 100 magnification was used to determine
the cercarial index, with the same oculars and photographic ocular as mentioned
above. The distances (in µm) were determined for 120 cercariae in each
isolate.
Determination of the number of dorsal and
ventral papillae - The papillae on the dorsal and ventral faces of the cercarial
body were counted and the papillae on the right and left hemi-bodies of the
two faces were quantified.
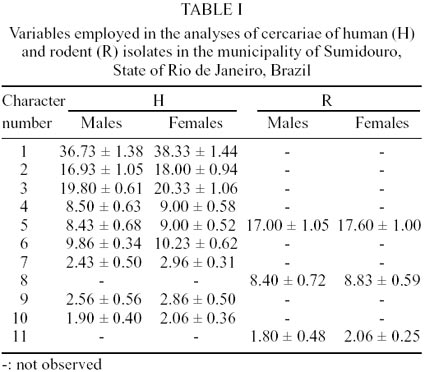
Spatial disposition of papillae - We used
a combination of the methodologies as proposed by Richard (1971) and Short and
Cartrett (1973) with slight modifications. The cercarial body was divided in
four parts: cephalic, pre-acetabular, medial e post-acetabular. The papillae
were numbered starting from the first cephalic papilla (Fig.
1C, D). The displacement or absence of papillae were recorded as was the
presence of supernumerary papillae, according to the number and position in
one of the segments of the cercarial body, on the right and left hemi-bodies
of the dorsal and ventral faces. For each isolate, at least 30 cercariae were
drawn.
Sex determination of cercariae - Mus
musculus Swiss Webster supplied by the Center for Laboratory Animal Breeding
at the Oswaldo Cruz Institute-Fiocruz, Brazil, were exposed to 50 cercariae
each shed by B. glabrata from the H or R isolates. Six animals were used
for each isolate. After six weeks, the animals were killed and perfused (Smithers
& Terry 1965) while the worms were identified according to their sex. The
animals were fed processed fodder for mice and rats (NUVILAB®)
ad libiditum, as well as water. The sacrifice of the animals followed the standards
in force regarding ethical procedures for laboratory animals (GV- SOLAS 1985).
Statistical analysis - Student's t test
was used for determination of mean differences of the chaetotaxic values and
the multivariate discriminating analysis (Morrison 1967) to distinguish the
observations on cercarial sex. Eleven variables were used (Table
I). The significance level adopted was 5%. The data processing was carried
out using BMDP-7M software.
RESULTS
Cercarial index - For the male cercariae
from the H isolate the cercarial index was 1.6 ± 0.8 µm and for
the R isolate 1.3 ± 0.3 µm. Among the females, the indices from
the R isolate (1.4 ± 0.3 µm) were greater than for the H isolate
(1.2 ± 0.3 µm). According to Student's t test, the values obtained
for males and females in both isolates were considered significant at the 5%
level. When a comparison between the indices presented by males and females
of each isolate were made, the difference was also significant at the 5% level.
Total number of dorsal and ventral papillae
- This varied from 38 to 46. Both isolates presented a greater variation on
the dorsal face (18-25) than on the ventral face (17-23).
Spatial disposition of papillae - In both
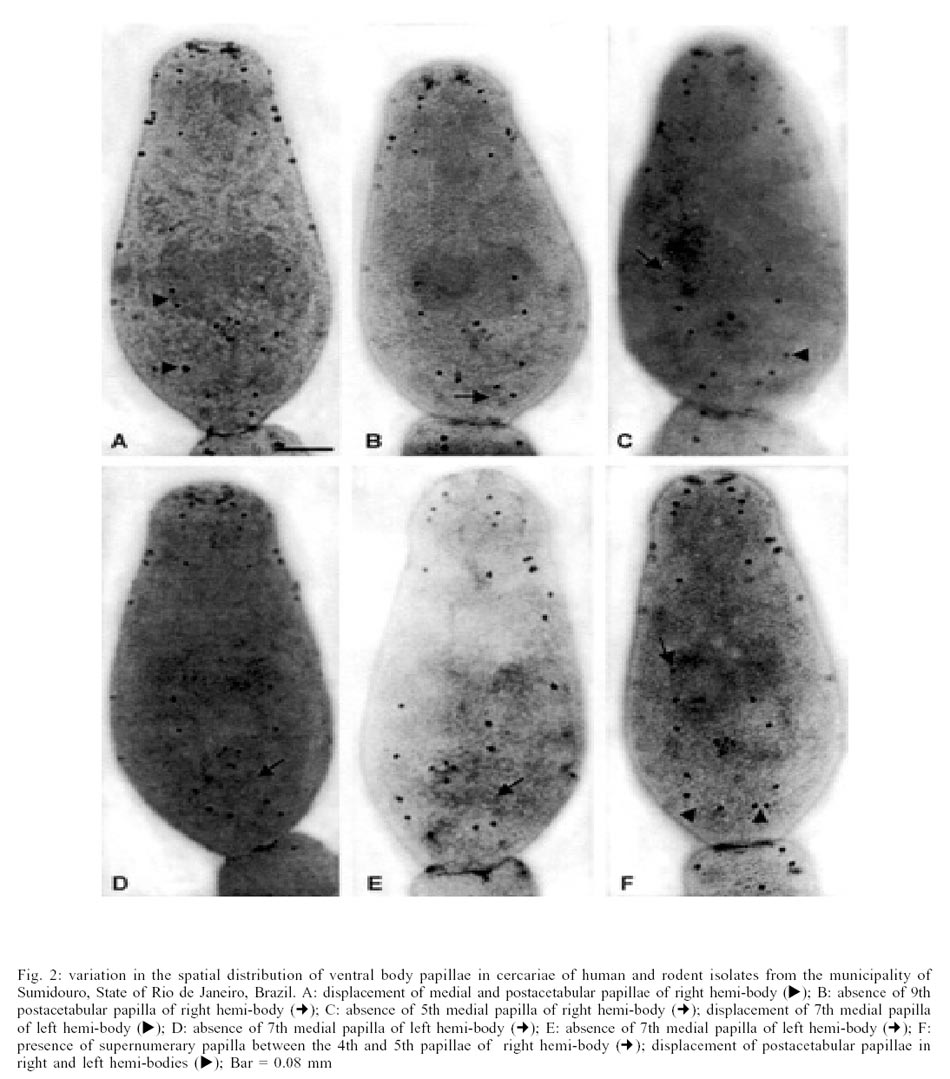
isolates a marked variability was observed on the ventral face of the cercariae.
On this face there was a marked constancy of papillae located on the cephalic
segment (V1, V2 and V3) and the preacetabular (V4) of the cercarial body. The
highest number of displacements (Fig. 2A, C,
F), papillary absences (Fig. 2B, D, E)
or supernumerary papillae (Fig. 2F) occurred
on the medial (V5, V6 and V7) and the postac-etabular (V9 and V10) segments.
The lack of V7 papilla was observed in both hemi-bodies (Fig.
2D, E).
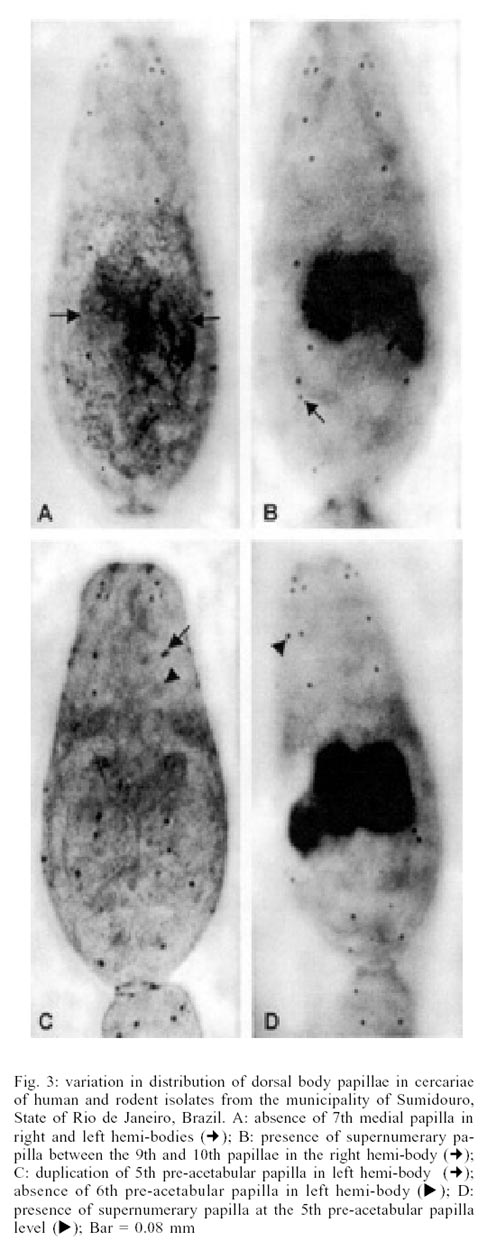
On the dorsal face, the greatest variability
occurred on the medial segment (papillae D7, D8 and D9). The cephalic segment
papillae (D1, D2, D3 and D4) and the preacetabular segment (D5 and D6) were
very constant. No variations in the acetabular papillae were observed with the
exception of a duplication of D5 on the left hemi-body (Fig.
3C). On this dorsal face there was also a lack of papillae (Fig.
3A, C) and supernumerary papillae (Fig.
3B, D).
All observations are valid for both isolates.
In relation to the sex of the cercariae, the greatest variability in the chaetotaxic
standard occurred in the males of both isolates.
None of the variables analyzed were able to distinguish
the isolates. In constrast, significant differences (at the 5% level) between
the sexes of the cercariae were observed. For isolate H, there were nine variables
and for isolate R three variables (Table II).
According to the Jackknifed classification, the variables that presented the
highest discriminatory power between the sexes were different for each isolate:
the number of papillae on the dorsal face of the right hemi-body (number 6),
the number of medial papillae on the ventral face of the right hemi-body (number
7) (isolate H) and the number of medial papillae on the dorsal face of the left
hemi-body (number 11) (isolate R).
DISCUSSION
Given that digenetic trematodes are primitively
snail parasites and not parasites of vertebrates (Richard 1971), it is convenient
to refer to the cercariae to base one's classification. The chaetotaxic study
of cercariae has been used as a tool to characterize strains of S. mansoni,
particularly where there are H and R strains (Bayssade-Dufour 1979b, Cassone
et al. 1981). However, in this trial with sympatric populations, a characteristic
cercarial index for each isolate was not observed, which points to the absence
of genetic variability between them. This observation is consistent with the
results obtained with sympatric isolates of H or murine origin from Senegal,
where the analysis of isoenzymes was used (Sène et al. 1997). It has
been demonstrated, through the use of DNA analysis, that strains that are geographically
close present homology (Pillay & Pillay 1994, Pinto et al. 1997). Even the
strains that are geographically different present over 90% similarity (Pillay
& Pillay 1994).
On the other hand, our data contrast with observations
that determined specific indices for strains from Africa and the Antilles with
different vertebrate hosts (non-human primate and rodent) (Bayssade-Dufour 1977).
This divergence is also present in the findings made with cercariae shed by
naturally infected snails and among H and R strains from Guadeloupe (Cassone
et al. 1981). In this case, indices around 1.30 could indicate the existence
of strains that moved alternately between H and R (Bayssade-Dufour 1979b). In
our opinion, this index, as well as other behavioral characteristics, present
a variation within certain limits. Therefore, what characterizes a certain isolate
is the frequency with which these values occur. The vertebrate hosts influence
this variability by selecting certain patterns. In this way, the cercarial index
may vary in the same location or in different locations, according to the participation
of each possible vertebrate host in the epidemiological chain of schistosomiasis
mansoni. In addition, this host can present subpopulations of worms with different
genotypes, as was shown in the R. rattus from Guadeloupe (Barral et al.
1996).
In our trial, no specific cercarial index was
observed for the different isolates. This constrast with demonstrations made
with other Brazilian strains (LE and SJ) (Araújo et al. 1987), which
are allopatric and have been kept for more than 30 years under laboratory conditions.
This condition is recognized as one of the factors that can lead to the selection
of a strain's chaetotaxic pattern (Cassone et al. 1978). It must be noted that
the isolates that were used in our trial were in their first passage through
laboratory conditions. It has been demonstrated that isolates recently introduced
to the laboratory present a greater genomic variability than those kept for
more generations (Vieira et al. 1991, Pinto et al. 1997).
With regard to the number and spatial distribution
of the body papillae, we observed that certain papillary clusters are more constant
than others and that the dorsal face is more stable than the ventral one. These
results confirm previous information regarding the existence of more or less
stable papillary clusters (Richard 1971, Bayssade-Dufour 1979a). We observed
a marked absence of the 7th ventral papilla (medial segment) in the Sumidouro
isolates. This fact was not recorded for any cercariae papillae of a Puerto
Rican strain (Short & Cartrett 1973).
The results of the present trial demonstrate
that the sex of the cercariae can be distinguished at the phenotypic level though
chromosomal analyses also allow sex determination (Quack et al. 1998). Furthermore,
differences in the chaetotaxic pattern of male and female cercariae have been
confirmed (Pino et al. 1988). Unlike these authors, however we observed that
the female cercariae presents greater homogeneity in the total number of papillae
of the cercarial body. Discriminant analysis applied to chaetotaxy is a fundamental
tool for separating cercariae of both sexes. Thus, about 66% cases were exactly
classified according to the total number of papillae (Pino & Morales, 1990).
In this experiment similar data were found. The smallest deviations in cercarial
indices, the smaller difference between the male and female indices and the
lower number of variables that are able to distinguish the sexes, indicate that
the R isolate is more homogenous than the H isolate. Morphological studies in
adult worms of these same isolates support this opinion (Machado-Silva et al.
1994, Neves et al. 1998). The results suggest that in this focus (Pamparrão
Valley, Sumidouro), R and H isolates belong to the same population that could
complete its life cycle in a rodent-snail-rodent fashion.
ACKNOWLEDGEMENTS
To Genilton José Oliveira and Heloísa
Maria Nogueira Diniz, Education Department, Oswaldo Cruz Institute, for their
technical support with the photographs. To Roberto Magalhães Pinto, Laboratory
of Helminths Parasites of Vertebrates, Helminthology Department, Oswaldo Cruz
Institute, for his collaboration with cercariae morphometry.
REFERENCES


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}