
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 2, 2002, pp. 143-150
|
Memórias do Instituto Oswaldo Cruz -
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(2) 2002, pp. 143-150
HIV-1 Polymorphism: a Challenge
for Vaccine Development - A Review
MG Morgado/+, ML Guimarães,
B Galvão-Castro*
Laboratório de AIDS e Imunologia
Molecular, Departamento de Imunologia, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil
4365, 21045-900 Rio de Janeiro, RJ, Brasil *Laboratório Avançado de Saúde Pública,
Centro de Pesquisa Gonçalo Moniz-Fiocruz, Salvador, BA, Brasil
+Corresponding author. Fax: +55-21-2280.1589. E-mail: mmorgado@ioc.fiocruz.br
Fiocruz is an UNAIDS Collaborative
Centre.
This work was supported by the National Coordination of STD/AIDS/UNDP/World
Bank, (UNAIDS), PIAF/Fiocruz/Ministry of Health, Faperj and Brazilian Research
Council (CNPq).
Received 5 January 2002
Accepted 15 February 2002
Code Number: oc02028
The perspective for the development
of anti-HIV/AIDS vaccines became a target sought by several research groups
and pharmaceutical companies. However, the complex virus biology in addition
to a striking genetic variability and the limited understanding of the immunological
correlates of protection have made this an enormous scientific challenge not
overcome so far. In this review we presented an updating of HIV-1 subtypes and
recombinant viruses circulating in South American countries, focusing mainly
on Brazil, as one of the challenges for HIV vaccine development. Moreover, we
discussed the importance of stimulating developing countries to participate
in the process of vaccine evaluation, not only testing vaccines according to
already defined protocols, but also working together with them, in order to
take into consideration their local information on virus diversity and host
genetic background relevant for the vaccine development and testing, as well
as including local virus based reagents to evaluate the immunogenicity of the
candidate vaccines.
Key words: HIV-1 polymorphism - HIV/AIDS
- vaccine
Since the identification of the viral
etiology of the Acquired Immunodeficiency Syndrome (AIDS) in 1983, with the
isolation of the Immunodeficiency Virus Type 1 (HIV-1) from patients with persistent
lymphadenopathy (Barre-Sinousi et al. 1983), the perspective for the development
of anti-HIV/AIDS vaccines became a target sought by several research groups
and pharmaceutical companies. However, the complex virus biology in addition
to a striking genetic variability and the limited understanding of the immunological
correlates of protection have made this an enormous scientific challenge not
overcome so far.
Indeed, HIV is one of the most important
emergent pathogens of this century. Based on phylogenetic analyses, it was estimated
to have emerged in the human population around 1931 (Korber et al. 2000), becoming
a worldwide public health challenge fifty years later.
As recently reviewed by the Joint
United Nations Programme on HIV/AIDS (UNAIDS 2001), since the description of
the first cases at the early beginning of the eighties in USA and Europe, more
than 21,8 million deaths have occurred due to HIV/AIDS. Forty million individuals,
adults and children, were estimated to be living with HIV/AIDS worldwide by
the end of 2001 and Latin America contributes with almost 4% of these cases.
HIV/AIDS was the cause of 4.2% of deaths in the world in 1999 being the fifth
cause of death worldwide. Only in 2001, 5.3 million adults and children were
estimated to be infected worldwide, with 14,000 new infections a day. Around
95% of these new cases occurred in developing countries. Roughly 14% were in
children under 15 and 86% in people aged 15-49, of whom almost 50% are women
and about 50% are 15-24 years old.
Although the development of combined
antiretroviral therapy has had a great impact on the reduction of AIDS morbidity
and mortality, its application is quite limited worldwide due to the high cost
and complex medical approach for the more deprived areas, complicated by the
emergence of drug resistant strains. Thus, the development of HIV/AIDS vaccine
is of high priority for public health.
There is a newly growing optimism
in the international scientific community concerning the possibility of developing
safe and effective preventive vaccines. However, despite the progress in recent
years, one of the major points is still to be clarified concerning the importance
of the intriguing virus diversity on vaccine efficacy. Indeed, viruses isolated
from different patients, and especially from different geographic regions, show
considerable genetic and thus antigenic variations, which may be a limiting
factor to the development of a universal vaccine.
MOLECULAR EPIDEMIOLOGY
OF HIV
The molecular epidemiology of HIV
and its potential implications for vaccine development and efficacy, as well
as for other aspects of HIV infection such as local epidemiological features,
virus transmission and pathogenesis, has been an issue of great concern (reviewed
by Workshop Report 1997a,b). Indeed, HIV is a highly polymorphic virus and two
types, HIV-1 and HIV-2, are participating in the AIDS epidemic. While HIV-2
has been basically restricted to the African continent, HIV-1 is associated
with the worldwide pandemic. Phylogenetic analyses of virus samples from different
geographic regions have revealed that HIV-1 can be divided into three groups:
O (outlier), N (new, non-N, non-O) and the M group (Major). While samples from
groups O and N are limited to the African continent, with some isolated cases
of group O in Europe and in the United States, group M variants are responsible
for the AIDS pandemic and were found to be subdivided into major genetic forms,
including distinct subtypes and sub-subtypes (A1, A2, B, C, D, F1, F2, G, H,
J and K). Moreover, 14 circulating recombinant forms (CRF 1 to 14), in addition
to unique recombinant viruses and several other HIV-1 unclassified genomes have
already been identified (reviewed in McCutchan 2000, CRF homepage http://hiv-web.lanl.gov/ for continuous
updating), as well as mixed infections (Janini et al. 1996, 1998, Ramos et al.
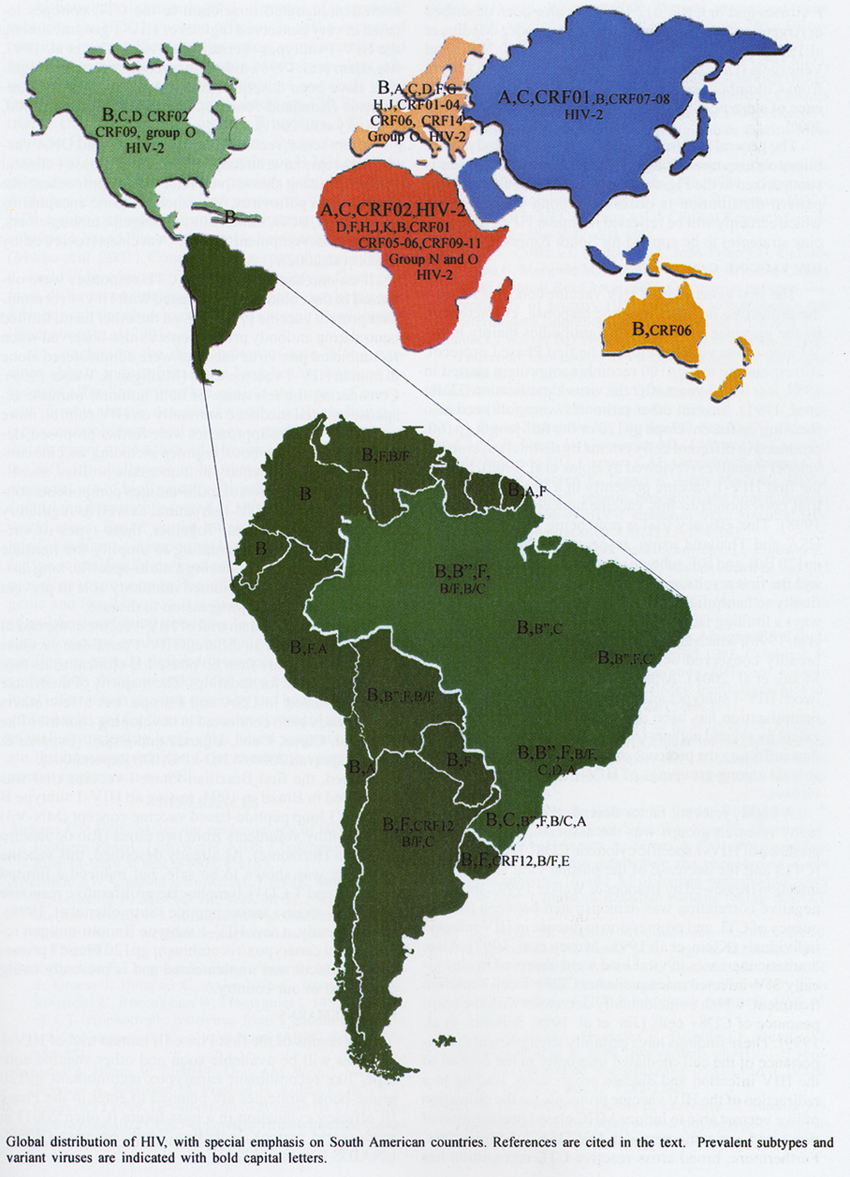
1999, Flores et al. 1999). This striking worldwide genetic variability is depicted
in the Figure.
The HIV-1 subtypes are geographically
distributed, with multiple subtypes and recombinant viruses co-circulating in
many areas of the world. All of them are similarly transmitted through sexual
contact, blood contamination, and vertically from mother to child.
Pairwise nucleotide distance among
the HIV-1 subtypes showed 20% for the envelope, 16% for the gag and 13% for
the polymerase genomic regions (Robertson et al. 1999). Such a high genetic
variability has direct influence on the antigenic constitution of these viruses.
Indeed, comparative analyses of relevant B cell, CD4+ T helper cell
and cytotoxic CD8+ T-lymphocytes (CTL) epitopes located in these
regions have shown an important level of variability in a greater or lesser
extent among the HIV-1 subtypes (HIV Molecular Immunology 2000).
It is therefore important to know
whether the HIV-1 strains circulating in the countries where the vaccine will
tentatively be used, are genetically and antigenically related to the vaccine
prototype being tested. In order to obtain this type of information, it is necessary
to keep HIV molecular surveillance programs, checking systematically the distribution
of the HIV-1 subtypes, as well as the introduction of new variants.
With the perspective of HIV/AIDS
preventive vaccine trials at the beginning of the nineties to be conducted in
different world regions, in both developed and developing countries, the knowledge
of the molecular epidemiology of HIV rapidly increased worldwide (Figure) and Brazil was one of the countries
participating in this international effort.
Several studies on the molecular
epidemiology of HIV-1 in Brazil showed the predominance of subtype B, with the
presence of additional subtypes like F (Morgado et al. 1994, 1996, 1998a, Galvão-Castro
et al. 1996, Tanuri et al. 1999, Brazilian Network for HIV Isolation and Characterization
2000), C (WHO 1994, Loureiro et al. 1998, Antunes 1998), D (Morgado et al. 1998a,
Tanuri et al. 1999) and, more recently, the presence of subtype A (Caride et
al. 2001, Guimarães et al. unpublished). Recombinant B/F and B/C viruses have
also been described (Sabino et al. 1994, Gao et al. 1996, Cornelissen et al.
1996, Tanuri et al. 1999, Vicente et al. 2000, Guimarães et al. 2001), in addition
to dual subtype infections (Janini et al 1996, 1998, Ramos et al. 1999).
Due to the interest for vaccine purposes,
the majority of these studies were originally performed based on the analysis
of the envelope gene, and, more recently, the evaluation of gag and polymerase
genomic polymorphisms have already been assessed. In addition to the inter-subtype
diversity, genetic and antigenic differences have also been described among
the subtype B viruses circulating in Brazil, with the identification of a subtype
B variant called B", which differs from the classical subtype B viruses by the
presence of the GWGR motif at the top of the envelope gp120 V3 loop, instead
of the GPGR molecular signature (Potts et al. 1993, Morgado et al. 1994, Brazilian
Network for HIV Isolation and Characterization 2000). These two subtype B variants
can be distinguished by both genetic (Morgado et al. 1998b, Covas et al. 1998)
and antigenic (Bongertz et al. 1994, Hendry et al. 1996, Casseb et al. 1998)
approaches, although consistent cross-neutralization has been observed between
them (Bongertz et al. 1997, 1998). Such observations are of important consideration
for vaccine evaluation in our country as the B" subtype B variant was found
to be highly prevalent in some Brazilian areas, corresponding to 57% of the
subtype B samples detected in Ribeirão Preto, SP (Covas et al. 1998) and 37%
of those from Rio de Janeiro, RJ (Morgado et al. 1998a,b).
Brazil is a huge country and differences
in the pattern of subtype distribution have been identified among the geographic
regions. Indeed, whereas similar proportions of B and F subtypes were observed
in a study conducted in Manaus (northern Brazil) (Vicente et al. 2000), in the
southeast, several studies have shown the predominance of subtype B (~85%) followed
by subtype F (~10-15%) (Morgado et al. 1994, 1998a, Tanuri et al. 1999, Brazilian
Network for HIV Isolation and Characterization 2000). Moreover, a clear predominance
of subtype B (> 90%), with very limited cases of F and C subtypes were, respectively,
observed in HIV-1 samples from the northeast (Couto-Fernandez et al. 1999) and
central-west (Stefani et al. 2000). The presence of subtype C in south Brazil
was firstly detected in 1 out 5 HIV-1 samples collected in Porto Alegre, RS,
in a setting of a WHO international study (WHO 1994). In a recent analysis conducted
by our group (unpublished data), we observed in 34% of the HIV-1 subtype C infected
samples collected in Porto Alegre, RS, in 1998, strongly contrasting with the
pattern of HIV-1 subtype distribution already verified for other Brazilian geographic
regions. This subtype seems to be spreading all over the world and is presently
the most prevalent strain in the AIDS pandemic (Esparza & Bhamarapravati
2000).
Following the Brazilian pattern,
studies on the molecular epidemiology of HIV-1 in other South American countries
also showed the predominance of subtype B, with the presence of additional subtypes
like F in Argentina, Bolivia, Uruguay, Peru, Paraguay, and Venezuela (Campodonico
et al. 1996, Marquina et al.1996, Velarde-Dunois et al. 2000, Masciotra et al.
2000, Russel et al. 2000, Castro et al. unpublished) and isolated cases of A
in Chile (Desgranges et al. 1998), Peru (Russel et al. 2000) and French Guyana
(Kazanji et al. 2001) and E in Uruguay (Artenstein et al. 1995). Moreover, unique
recombinant B/F viruses and/or CRF_012 B/F have also been described in Argentina
(Marquina et al. 1996, Fernandez-Medina et al.1999, Carr et al. 2001), Uruguay
(Carr et al. 2001) and Venezuela (Delgado et al. 2001, Castro et al. 2001).
Data from Colombia and Ecuador have only shown the presence of subtype B so
far (Navas et al. 1999, Russel et al. 2000).
The general picture of the HIV-1
subtypes and recombinant viruses circulating in South American countries is
summarized in the Figure. In general, a quite similar virus
pattern distribution is observed among the countries, which certainly will be
reflected in similar HIV/AIDS vaccine strategies to be applied for South America.
HIV VACCINE CANDIDATES
The first generation of HIV vaccine
concepts favored the protective humoral immune response, characterized by the
presence of neutralizing antibodies mainly to the V3 loop of the viral envelope.
The first Phase I protocol, corresponding to a gp160 recombinant protein started
in 1987, less than 5 years after the virus identification (Dolin et al. 1991).
Several other protocols were followed also focusing on the envelope gp120 or
the full-length gp160, produced in different cell systems by distinct vaccine
developer initiatives (reviewed by Bojak et al. 2002). Indeed, the first HIV-1
vaccine presently in a Phase III efficacy trial corresponds to this vaccine
concept (Francis et al. 1998). This efficacy trial is now being conducted in
the USA and Thailand using, respectively, bivalent HIV-1 gp120 B/B and B/E subtype
vaccines (Berman et al. 1999), and the first results are expected soon. However,
the difficulty to neutralize primary isolates of the virus was always a limiting
factor of these vaccine concepts (Mascola et al. 1996), which are not able to
induce antibodies against broadly conserved neutralizing epitopes (reviewed
by Moore et al. 2001). Although there is no correlation between HIV-1 subtypes
and virus neutralization, cross-clade neutralization has been described in a
greater or lesser extent by several authors (reviewed by Moore et al. 2001),
thus reflecting the presence of common antibody epitopes, at least among groupings
of HIV-1 subtypes and variant viruses.
A highly relevant factor described
in recent years by many research groups was the association between the presence
of HIV-1 specific cytotoxic CD8+ T-lymphocytes (CTL) and the decrease
of the plasma viral load in HIV infection (reviewed by Brander & Walker
1999). Indeed, a negative correlation was demonstrated between the frequency
of CTL and progression to disease in HIV infected individuals (Klein et al.
1995, Musey et al. 1997). Also, dramatic increases in viral load were observed
in chronically SIV-infected macaques after CD8+ T cell depletion treatment,
which coincidentally decreases with the reappearance of CD8+ cells (Jin et al.
1999, Schmitz et al. 1999). These findings have certainly strengthened the importance
of the cell-mediated immunity in the control of the HIV infection and disease
progression, leading to a redirection of the HIV vaccine protocols for the utilization
of live vectors able to induce MHC class I presentation of processed antigenic
peptides to CD8+ T-lymphocytes. Furthermore, broad cross-reactive CTL recognition
has been demonstrated in relation to the CTL epitopes located in very conserved
regions of HIV-1 genome among the HIV-1 subtypes (Ferrari et al. 1997, Cao et
al. 1997, McAdam et al. 1998), although reactions of higher magnitude have been
described for subtype-specific CTL responses (Rowland-Jones et al. 1998, Cao
et al. 2000, Novitsky et al. 2001).
In this sense, recombinant pox viruses
and DNA vaccine concepts have already been tested in Phase I clinical trials.
Promising alternative vector-based approaches, like recombinant poliovirus,
Venezuelan equine encephalitis virus (VEE), BCG, salmonella and shigella, among
others, are under development for HIV vaccines (reviewed by Bojak et al. 2002).
If on one hand, no specific CTL responses
were observed in the volunteers vaccinated with HIV-1 recombinant protein vaccine
protocols, on the other hand, limited neutralizing antibody production was also
observed when recombinant pox virus vaccines were administered alone in human
HIV-1 vaccine trials (Mulligan & Weber 1999). Considering the relevance
of both humoral immune response and cell mediated immunity on HIV control, more
promising vaccine approaches were further proposed, denominated as prime-boost
regimen including vaccine concepts with complementary immunogenic profiles,
associated with the inclusion of multi-antigen components consisting of selected
HIV-1 structural as well as regulatory antigen-encoding genes. Together, these
types of vaccine approaches are attempting to amplify the immune response to
the virus, inducing a multi-specific, long lasting humoral and cell mediated
immunity able to prevent HIV infection and/or progression to disease.
Since the first human trial of HIV-vaccine
at the end of the eighties, around 30 different HIV-1 candidate vaccines were
tested in more than 60 phase I/II clinical trials (see Bojak et al. 2002 for
updating). The majority of them have been conducted in USA and Europe, but a
few others have already been conducted in developing countries like Thailand,
China, Cuba, Uganda and Brazil (Esparza & Bhamarapravati 2000, WHO-UNAIDS
Report 2001).
Indeed, the first Brazilian Phase
I vaccine trial was conducted in Brazil in 1994, testing an HIV-1 subtype B
gp120 V3 loop peptide-based vaccine concept (MN-V3) in 30 healthy volunteers
from two cities (Rio de Janeiro and Belo Horizonte). As already described, this
vaccine candidate was shown to be safe, but induced a limited antibody and T
CD4+ lymphocyte proliferative response to the MN strain vaccine peptide (Sutmoller
et al. 1998). More recently, a new HIV-1 subtype B multi-antigen recombinant
canarypox/recombinant gp120 Phase I prime-boost protocol was implemented and
is presently being conducted in our country.
FINAL REMARKS
The results of the first Phase III
human trial of HIV-1 vaccines will be available soon and other vaccine concepts
like recombinant canarypox/recombinant gp120 prime-boost strategies are planned
to enter in the Phase III efficacy evaluation in a near future (Cohen 2001)
in some world regions, including South America (WHO-UNAIDS Report 2001).
Although the immunological relevance
of the genetic subtypes for HIV-1 vaccine efficacy is still a matter of extensive
investigation and discussion, some international efforts were made for the inclusion
of local prevalent subtypes and variant viruses in the vaccine protocols to
be tested. For instance, the recombinant gp 120 Phase III trial in USA includes
two subtype B variants, MN and a subtype B primary isolate, whereas the protocol
being conducted in Thailand employs subtype B MN and a region-specific gp120
subtype E primary isolate (Mulligan & Weber 1999). In the African continent,
where several HIV-1 subtypes and recombinant viruses predominate, a subtype
A based Phase I protocol is being tested in Kenya (Mwau et al. 2001). Considering
the spreading of subtype C worldwide, comparative analysis of subtype C samples
from different countries, including Brazil, are being performed with the objective
of future vaccine development (Rodenburg et al. 2001).
In conclusion, the development of
safe HIV-1 vaccines able to induce broad and long-lasting immune responses affording
protection against the array of prevalent variants already described worldwide
(including subtypes, recombinants and variant viruses) is presently a distant
target to be achieved. However, even considering the difficult challenges to
be overcome, urgent vaccine measures are of high priority for public health
in order to control the devastating spread of HIV mainly in the poorer world
areas. Based on the knowledge already accumulated over time, it is important
to stimulate Phase I/II trials for the scientifically defined promising vaccine
concepts as well as to move on to Phase III with the more immunogenic and feasible
ones. It is of paramount importance to stimulate developing countries to participate
in the process of vaccine evaluation, not only testing vaccines according to
already defined protocols. But, working together with them, in order take into
consideration their local information on virus diversity and host genetic background
relevant for the vaccine development and testing, as well as including local
virus based reagents to evaluate the immunogenicity of the candidate vaccines
is of utmost importance as well.
REFERENCES
- Antunes CV 1998. Molecular epidemiology of HIV/AIDS in southern Brazil.
12th World AIDS Conference, Geneva, June 28- July 3, abst. 61003, p.1183.
- Artenstein AW, Coppola J, Brown AE, Carr JK, Sanders-Buell E, Galbarini
E, Mascola JR, VanCott TC, Schonbrood P, McCutchan FE 1995. Multiple introductions
of HIV-1 subtype E into the western hemisphere. Lancet 346 (8984):
1197-1198.
- Barré-Sinoussi F, Cherman JC, Rey F, Nugeyre MT, Chamaret S, Gruest J, Dauguet
C, Axler-Blin C, Brun-Vezinet F, Rouzioux C, Rozenbaum W, Montagnier L 1983.
Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired
immune deficiency syndrome (AIDS). Science 220: 868-871.
- Berman PW, Huang W, Riddle L, Gray AM, Wrin T, Vennari J, Johnson A, Klaussen
M, Prashad H, Kohne C, deWit C, Gregory TJ 1999. Development of bivalent (B/E)
vaccines able to neutralize CCR5-dependent viruses from the United States
and Thailand. Virology 265:1-9.
- Bojak A, Deml L, Wagner R 2002. The past, present and future of HIV-vaccine
development: a critical view. Drug Discov Today 7: 36-46.
- Bongertz V, Costa CI, Guimarães ML, Soares da Costa MFG, Grinsztejn B, HEC/Fiocruz
AIDS Clinical Research Group, Bastos FI, Pilotto JH, João Filho EC, Loureiro
R, Chequer P, Telles PR, Galvão-Castro B, Morgado MG 1998a. HIV specific humoral
immune response in Rio de Janeiro. Mem Inst Oswaldo Cruz 93: 391-398.
- Bongertz V, Costa CI, Guimarães ML, Grinsztejn B, João Filho EC, Galvão-Castro
B, Morgado MG 1998b. Neutralization susceptibility of B subtype variant B"
primary HIV-1 isolates. The HEC/Fiocruz AIDS Clinical Research Group. Scand
J Immunol 47: 603-608.
- Bongertz V, Costa CI, Santos VGV, João Filho EC, Galvão-Castro B, Morgado
MG, and the Hospital Evandro Chagas AIDS Clinical Res Group 1997. Correlation
between susceptibility of primary HIV-1 isolates to autologous and heterologous
neutralizing antibodies. AIDS 11: 969-975.
- Brander C, Walker BD 1999. T lymphocyte responses in HIV-1 infection: implications
for vaccine development. Curr Opin Immunol 11: 451-459.
- Brazilian Network for the HIV-1 Isolation and Characterization (Bongertz
V, Bou-Habib DC, Brígido LFM, Caseiro M, Couto-Fernandez JC, Ferreira PC,
Freitas CO, Galvão-Castro B, Greco D, Guimarães ML, Linhares de Carvalho MI,
Morgado MG, Oliveira CAF, Osmanov S, Ramos CA, Rossini M, Sabino E, Tanuri
A, Ueda M) 2000. HIV-1 diversity in Brazil: genetic, biological and immunological
characterization of HIV-1 strains in three potential HIV vaccine evaluation
sites. J AIDS 23: 184-193.
- Campodonico M, Janssens W, Heyndrickx L, Fransen K, Leonaers A, Fay FF,
Taborda M, Van der Groen G, Fay OH 1996. HIV type 1 subtypes in Argentina
and genetic heterogeneity of the V3 region. AIDS Res Hum Retroviruses 12:
79-81.
- Cao H, Kanki P, Sankalé J-L, Dieng-Sarr A, Mazzara GP, Kalams SA, Korber
B, Mboup S, Walker BD 1997. Cytotoxic T-lymphocyte cross-reativity among different
Human Immudefinciency virus type 1 clades: Implications for vaccine development.
J Virol 71: 8615-8623.
- Cao H, Mani I, Vincent R, Mugerwa R, Mugyenyi P, Kanki P, Ellner J, Walker
BD 2000. Cellular immunity to human immunodeficiency virus type 1 (HIV-1)
clades: relevance to HIV-1 vaccine trials in Uganda. J Infect Dis 182:
1350-1356.
- Caride E, Brindeiro R, Hertogs K, Larder B, Dehertogh P, Machado E, Sá CA,
Eyer-Silva WA, Sion FS, Passioni LF, Menezes JA, Calazans A, Tanuri A 2001.
Drug-resistant reverse transcriptase genotyping and phenotyping B and non-B
subtypes (F and A) of human immunodeficiency virus type 1 found in Brazilian
patients falling HAART. Virology 275: 107-115.
- Carr JK, Avila M, Carrillo MG, Salomon H, Hierholzer J, Watanaveeradej V,
Pando MA, Negrete M, Russell KL, Sanchez J, Birx DL, Andrade R, Vinoles J,
McCutchan FE 2001. Diverse BF recombinants have spread widely since the introduction
of HIV-1 into South America. AIDS 15: F41-F47.
- Casseb J, Hong MA, Gonsalez C, Brigido LF, Duarte AJ, Michael-Hendry R 1998.Two
variants of HIV-1 B serotype are transmitted heterosexually in São Paulo,
Brazil. Braz J Med Biol Res 31: 1243-1246.
- Castro E, Echeverría G, Deibis L, Gonzales B, Moreira AS, Morgado MG 2001.
Subtype and phylogenetics analysis of HIV-1 isolates from Venezuela. The first
IAS Conference on HIV pathogeneses and treatments, Buenos Aires, Argentina,
July 2001.
- Cohen J 2001. AIDS research. Debate begins over new vaccine trials. Science
293(5537): 1973.
- Cornelissen M, Kampinga G, Zorgdrager F, Goudsmit J, the UNAIDS network
for HIV isolation and characterization 1996. Human immnodeficiency virus type
1 subtypes defined by env show high frequency of recombination gag genes.
J Virol 70: 8209-8212.
- Couto-Fernandez JC, Morgado MG, Bongertz V, Tanuri A, Andrade T, Brites
C, Galvão-Castro B 1999. HIV-1 subtyping in Salvador, Bahia, Brazil: a city
with African sociodemographic characteristics. J Acquir Immune Defic Syndr
22: 288-2893.
- Covas DT, Biscaro TA, Kashima S, Duarte G, Machado AA 1998. High frequency
of the GWG (Pro Trp) envelope variant of HIV-1 in Southeast Brazil. J Acquir
Immune Defic Syndr Hum Retrovirol 19: 74-99.
- CRF homepage – http://hiv-web.lanl.gov/seq-db.html
- Delgado E, Leon-Ponte M, Villahermosa ML, Cuevas MT, Deibis L, Echeverria
G, Thomson MM, Perez-Alvarez L, Osmanov S, Najera R 2001. Analysis of HIV
type 1 protease and reverse transcriptase sequences from Venezuela for drug
resistance-associated mutations and subtype classification: a UNAIDS study.
AIDS Res Hum Retroviruses 17: 753-758.
- Desgranges C, Fillon S, Letouneur F, Buzelay L, Sepulveda C, Guzman M-A,
Afani A, Barin F, Saragosti S 1998. HIV-1 subtypes in Santiago, Chile. AIDS
12: 1563-1565.
- Dolin R, Graham BS, Greenberg SB, Tacket CO, Belshe RB, Midthun K, Clements
ML, Gorse GJ, Horgan BW, Atmar RL 1991. The safety and immunogenicity of a
human immunodeficiency virus type 1 (HIV-1) recombinant gp160 candidate vaccine
in humans. NIAID AIDS Vaccine Clinical Trials Network. Ann Intern Med 114:
119-127.
- Esparza J, Bhamarapravati N 2000. Accelerating the development and future
availability of HIV-1 vaccines: Why, when, where and how? The Lancet 355:
2061-2066.
- Fernandez-Medina D, Jansson M, Rabinovich RD, Libonatti O, Wigzell H 1999.
Identification of human immunodeficiency virus type 1 subtypes B and F B/F
recombinant and dual infection with these subtypes in Argentina. Scand
J Infect Dis 31: 235-242.
- Ferrari G, Humphrey W, McElrath MJ, Excler JL, Duliege AM, Clements ML,
Corey LC, Bolognesi DP, Weinhold KJ 1997. Clade B-based HIV-1 vaccines elicit
cross-clade cytotoxic T lymphocyte reactivities in uninfected volunteers.
Proc Natl Acad Sci USA 94: 1396-1401.
- Flores I, Pieniazek D, Moran N, Soler A, Rodriguez N, Alegria M, Vera M,
Janini LM, Bandea CI, Ramos A, Rayfield M, Yamamura Y 1999. HIV-1 subtype
F and dual infections in Puerto Rico: a potential sentinel site for monitoring
novel genetic HIV variants in North America. Emerg Infect Dis 5: 481-483.
- Francis DP, Gregory T, McElrath MJ, Belshe RB, Gorse GJ, Migasena S, Kitayaporn
D, Pitisuttitham P, Matthews T, Schwartz DH, Berman PW 1998. Advancing AIDSVAX
to phase 3. Safety, immunogenicity, and plans for phase 3. AIDS Res Hum
Retroviruses 14 (Suppl. 3): S325-331.
- Galvão-Castro B, Couto-Fernandez JC, Mello MA, Linhares-de-Carvalho MI,
Castello-Branco LR, Bongertz V, Ferreira PC, Morgado M, Sabino E, Tanuri A
1996. A nationwide effort to sistematically monitor HIV-1 diversity in Brazil:
preliminary results. Brazilian Network for the HIV-1 Isolation and Characterization.
Mem Inst Oswaldo Cruz 91: 335-338.
- Gao F, Robertson DL, Morrison SG, Hui H, Craig S, Decker J, Fultz PN, Girard
M, Shaw GM, Hahn BH, Sharp PM 1996. The heterosexual human immunodeficiency
virus type 1 epidemic in Thailand is caused by an intersubtype (A/E) recombinant
of African origin. J Virol 70: 7013-7029.
- Graham BS 2000. Los Alamos HIV Molecular Immunology Database 2000. In BTM
Korber, C Brander, BF Haynes, R Koup, C Kuiken, JP Moore, BD Walker, D Watkins
(eds), Los Alamos National Laboratory: Theoretical Biology and Biophysics,
Los Alamos, NM.
- Guimarães M, Moreira A, Morgado M 2001. Molecular characterization
of CTL epitopes among prevalent Brazilian HIV-1 subtypes. AIDS Vaccine 2001,
Philadelphia, PA, USA, abstract 261.
- Hendry RM, Hanson CV, Bongertz V, Morgado M, Duarte A, Casseb J, Brígido
L, Sabino E, Diaz R, Galvão-Castro B 1996. Immunoreactivity of Brazilian HIV
isolates with different V3 motifs. Mem Inst Oswaldo Cruz 91: 347-348.
- HIV Molecular Immunology Database 2000. In BTM Korber, C Brander, BF Haynes,
R Koup, C Kuiken, JP Moore, BD Walker, D Watkins (eds), Los Alamos National
Laboratory: Theoretical Biology and Biophysics, Los Alamos, NM.
- Janini LM, Pieniazek D, Peralta JM, Schechter M, Tanuri A, Vicente AC, dela
Torre N, Pieniazek NJ, Luo CC, Kalish ML, Schochetman G, Rayfield MA 1996.
Identification of single and dual infections with distinct subtypes of human
immunodeficiency virus type 1 by using restriction fragment length polymorphism
analysis. Virus Genes 13: 69-81.
- Janini LM, Tanuri A, Schechter M, Peralta JM, Vicente AC, Dela Torre N,
Pieniazek NJ, Luo CC, Ramos A, Soriano V, Schochetman G, Rayfield MA, Pieniazek
D 1998. Horizontal and vertical transmission of human immunodeficiency virus
type 1 dual infections caused by viruses of subtypes B and C. J Infect
Dis 177: 227-231.
- Jin X, Bauer DE, Tuttleton SE, Lewin S, Gettie A, Blanchard J, Irwin CE,
Safrit JT, Mittler J, Weinberger L, Kostrikis LG, Zhang L, Perelson AS, Ho
DD 1999. Dramatic rise in plasma viremia after CD8(+) T cell depletion in
simian immunodeficiency virus-infected macaques. J Exp Med 189 : 991-998.
- Kazanji M, Lavergne A, Pouliquen J-F, Magnien C, Bissuel F, Marty C, Couppie
P, Talarmin A 2001. Gemetic diversity and phylogenetic analysis of human immunodeficiency
virus type 1 subtypes circulating in French Guiana. AIDS Res Hum Retrovirus
17: 857-861.
- Klein MR, van Baalen CA, Holwerda AM, Kerkhof Garde SR, Bende RJ, Keet IP,
Eeftinck-Schattenkerk JK, Osterhaus AD, Schuitemaker H, Miedema F 1995. Kinetics
of Gag-specific cytotoxic T lymphocyte responses during the clinical course
of HIV-1 infection: a longitudinal analysis of rapid progressors and long-term
asymptomatics. J Exp Med 181: 1365-1372.
- Korber B, Muldoon M, Theiler J, Gao F, Gupta R, Lapedes A, Hahn BH, Wolinsky
S, Bhattacharya T 2000. Timing the ancestor of the HIV-1 pandemic strains.
Science 288 (5472): 1789-1796.
- Loureiro R, Casseb JS, Bongertz V, Asquidanini S, Bet E, Silveira VLB, Dias
CS 1998. Seroreactivity to HIV-1 peptides in pregnant women in Porto Alegre,
RS State, Brazil. 12th World AIDS Conference, Geneva, June 28-July 3. Conference
Record, p. 22.
- Marquina S, Leitner T, Rabinovich RD, Benetucci, Libonatti O, Albert J 1996.
Coexistence of subtype B, F and B/F env recombinant of HIV type 1 in Buenos
Aires, Argentina. AIDS Res Hum Retroviruses 12: 1651-1654.
- Masciotra S, Livellara B, Belloso W, Clara L, Tanuri A, Ramos AC, Baggs
J, Lal R, Pieniazek D 2000. Evidence of a high frequency of HIV-1 subtype
F infections in a heterosexual population in Buenos Aires, Argentina. AIDS
Res Hum Retroviruses 16: 1007-1014.
- Mascola JR, Louder MK, Surman SR, Vancott TC, Yu XF, Bradac J, Porter KR,
Nelson KE, Girard M, McNeil JG, McCutchan FE, Birx DL, Burke DS 1996. Human
immunodeficiency virus type 1 neutralizing antibody serotyping using serum
pools and an infectivity reduction assay. AIDS Res Hum Retroviruses 12:
1319-1328.
- McAdam S, Kaleebu P, Krausa P, Goulder P, French N, Collin B, Blanchard
T, Whitworth J, McMichael A, Gotch F 1998. Cross-clade recognition of p55
by cytotoxic T lymphocytes in HIV-1 infection. AIDS 12: 571-579.
- McCutchan FE 2000. Understanding the genetic diversity of HIV-1. AIDS
14 (Suppl. 3): S31-S44.
- Moore JP, Parren PW, Burton DR 2001. Genetic subtypes, humoral immunity,
and human immunodeficiency virus type 1 vaccine development. J Virol 75:
5721-5729.
- Morgado MG, Guimarães ML, Gripp CBG, Neves Jr. I, Costa CI, Santos VGV,
Linhares-de-Carvalho MI, Galvão-Castro B, Bongertz V, The Hospital Evandro
Chagas AIDS Clinical Research Group 1998a. High prevalence of HIV-1 subtype
B and identification of an HIV-1 subtype D infection in the City of Rio de
Janeiro, Brazil. J AIDS Hum Retroviruses 18: 488-494.
- Morgado MG, Guimarães ML, Neves Jr I, Costa CI, Santos VGV, Linhares-de-Carvalho
MI, Galvão-Castro B, Bongertz V 1996. Polymorphism of the predictive antigenic
sites on the V3 loop of Brazilian HIV-1 subtype strains. Mem Inst Oswaldo
Cruz 91: 339-342.
- Morgado MG, Guimarães ML, Neves Jr I, Veloso VG, Linhares-de-Carvalho MI,
Castello-Branco LR, Bastos FI, Castilho EA, Galvão-Castro B, Bongertz V, The
Hospital Evandro Chagas AIDS Clinical Research Group 1998b. Polymorphism of
the antigenically distinct HIV-1 B subtype strains. Mem Inst Oswaldo Cruz
93: 383-386.
- Morgado MG, Sabino EC, Shpaer EG, Bongertz V, Brigido L, Guimarães MDC,
Castilho EA, Galvão-Castro B, Mullins JL, Hendry RM, Mayer A 1994. V3 region
polymorphism in HIV-1 from Brazil: Prevalence of subtype B strains divergent
from the North American/European prototype and detection of subtype F. AIDS
Res Hum Retroviruses 10: 569-575.
- Mulligan MJ, Weber J 1999. Human trials of HIV-1 vaccines. AIDS 13
(Suppl. A): S105-112.
- Musey L, Hughes J, Schacker T, Shea T, Corey L, McElrath MJ 1997. Cytotoxic-T-cell
responses, viral load, and disease progression in early human immunodeficiency
virus type 1 infection. N Engl J Med 337: 1267-1274.
- Mwau M, Patel S, Sutton J, Tomlinson M, Hanke T, McMichael A 2001. Clinical
evaluation of an HIV-1 clade A DNA/MVA vaccine designed for Kenya. AIDS Vaccine
2001, Philadelphia, PA, USA abstract S5.
- Navas M-A, Letourneur F, Gomas E, Boshell J, Saragosti S 1999. Analysis
of the V3 loop sequences from 12 HIV type 1-infected patients from Colombia,
South America. AIDS Res Hum Retroviruses 15: 1141-1144.
- Novitsky V, Rybak N, McLane MF, Gilbert P, Chigwedere P, Klein I, Gaolekwe
S, Chang SY, Peter T, Thior I, Ndung'u T, Vannberg F, Foley BT, Marlink R,
Lee TH, Essex M 2001. Identification of human immunodeficiency virus type
1 subtype C Gag-, Tat-, Rev-, and Nef-specific elispot-based cytotoxic T-lymphocyte
responses for AIDS vaccine design. J Virol 75: 9210-9228.
- Potts KE, Kalish ML, Lott T, Orloff G, Luo CC, Bernard MA, Alves CB, Badaro
R, Suleiman J, Ferreira O, and the Brazilian Collaborative AIDS Research Group
1993. Genetic heterogeneity of the V3 region of the HIV-1 envelope glycoprotein
in Brazil. AIDS 7: 1191-1197.
- Ramos A, Tanuri A, Schechter M, Rayfiel MA, Hu DJ, Cabral MC, Bandea CI,
Baggs J, Pieniazek D 1999. Dual and recombinant infections: an integral part
of the HIV-1 epidemic in Brazil. Emerg Infect Dis 5: 65-74.
- Robertson DL, Anderson JP, Bradac JA, Carr JK, Foley B, Funkhouser RK, Gao
F, Hahn BH, Kalish ML, Kuiken C, Learn GH, Leitner T, McCutchan F, Osmanov
S, Peeters M, Pieniazek D, Salminen M, Sharp PM, Wolinsky S, Korber B 1999.
Human Retroviruses and AIDS 1999. A Compilation and Analysis of Nucleic
Acid and Amino Acid Sequences, Los Alamos National Laboratory, Los Alamos,
NM.
- Rodenburg CM, Li Yingying, Trask SA, Chen Y, Decker J, Robertson DL, Kalish
ML, Shaw GM, Allen S, Hahn BH, Gao F and UNAIDS and NIAID Networks for HIV
isolation and characterization 2001. Near full-length clones and reference
sequences for subtype C isolates of HIV type 1 from three different continents.
AIDS Res Hum Retroviruses 17: 161-168.
- Rowland-Jones SL, Dong T, Fowke KR, Kimani J, Krausa P, Newell H, Blanchard
T, Ariyoshi K, Oyugi J, Ngugi E, Bwayo J, MacDonald KS, McMichael AJ, Plummer
FA 1998. Cytotoxic T cell responses to multiple conserved HIV epitopes in
HIV-resistant prostitutes in Nairobi. J Clin Invest 102:1758-1765.
- Russel KL, Carcamo C, Watts DM, Sanchez J, Gotuzzo E, Euler A, Blanco JC,
Galeano A, Alava A, Mullins JI, Holmes KK, Carr JK 2000. Emerging genetic
diversity of HIV-1 in South America. AIDS 14: 1785-1791.
- Sabino E, Shpaer E, Morgado MG, Kober BT, Diaz RS, Bongertz V, Cavalcante
S, Galvão-Castro B, Mullins JI, Mayer A 1994. Identification of an HIV-1 proviral
genome recombinant between subtype B and F in PBMCs obtained from an individual
in Brazil. J Virol 68: 6340-6346.
- Schmitz JE, Kuroda MJ, Santra S, Sasseville VG, Simon MA, Lifton MA, Racz
P, Tenner-Racz K, Dalesandro M, Scallon BJ, Ghrayeb J, Forman MA, Montefiori
DC, Rieber EP, Letvin NL, Reimann KA 1999. Control of viremia in simian immunodeficiency
virus infection by CD8+ lymphocytes. Science 283(5403): 857-860.
- Stefani MM, Pereira GA, Martelli CM, Shindo N, Galvão-Castro B 2000. Evidence
of HIV-1 genetic diversity among pregnant women with AIDS or infected with
HIV-1 in Central Brazil. J Acquir Immune Defic Syndr 23: 205-207.
- Sutmoller F, Greco D, Morgado MG, Toledo Jr AC, Tupinambás U, Gripp CBWG,
Koff W for the Vaccin Protocol Team 1998. Safety and immunogenicity trial
of na HIV-1 MN V3 peptide vaccine in Brazil. International Meeting on Vaccines.
Salvador, BA, Brasil, p.102, abstract P15.213.
- Tanuri A, Swanson P, Devare S, Berro O, Savedra A, Costa LJ, Telles JG,
Brindeiro R, Schable C, Pieniazek D, Rayfield M 1999. HIV-1 subtypes among
blood donors from Rio de Janeiro, Brazil. J Acq Immune Defic Syndr Hum
Retr 20: 60-66.
- UNAIDS 2001- UNAIDS webpage. http://www.unaids.org/
- Velarde-Dunois KG, Guimarães M L, La Fuente C, Andrade R, Arévalo R, Pantoja
S, Mariscal R, Sandoval R, Iriarte F, Chamón V, Melgar M, Carvajal
R, Morgado MG 2000. Molecular characterization of human immunodeficiency virus
type-1 infected individuals from Bolivia reveals the presence of two distinct
genetic subtypes B and F. AIDS Res Hum Retroviruses 16: 1921-1926.
- Vicente ACP, Otsuki K, Silva NB, Castilho MC, Barros FS, Pieniazek D, Hu
D, Rayfield MA, Bretas G, Tanuri A 2000. The HIV epidemic in the Amazon basin
is driven by prototypic and recombinant HIV-1 subtypes B and F. J AIDS
23: 327-331.
- WHO Network for HIV Isolation and Characterization 1994. HIV-1 type 1 variation
in World Health Organization-sponsored vaccine evaluation sites: Genetic screening,
sequence analysis and preliminary biological characterization of selected
viral strains. AIDS Res Hum Retroviruses 10: 1327-1343.
- WHO/UNAIDS Report 2001. Future access to HIV vaccines. AIDS 15: W27-44.
- Wokshop Report from the European Commission (DG XII, ICNO-DC), Joint United
Nations Programme on HIV/AIDS 1997a. HIV-1 subtypes: implications for epidemiology,
pathogenicity, vaccines and diagnostics. AIDS 11: UNAIDS 17-36.
- Wokshop Report from the Expert Group of the Joint Nations Programme on HIV/HIDS
1997b. Implications of HIV variability for transmission: scientific and policy
issues. AIDS 11: UNAIDS 15.
© 2002
Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02028f1.jpg]
|

{kind=link}