 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(2) 2002, pp. 169-174 Cotton Rats (Sigmodon hispidus) and Black Rats (Rattus rattus) as Possible Reservoirs of Leishmania spp. in Lara State, Venezuela Hector De Lima, Zoraya De Guglielmo, Armando Rodríguez, Jacinto Convit, Noris Rodriguez+ Instituto de Biomedicina, Esquina
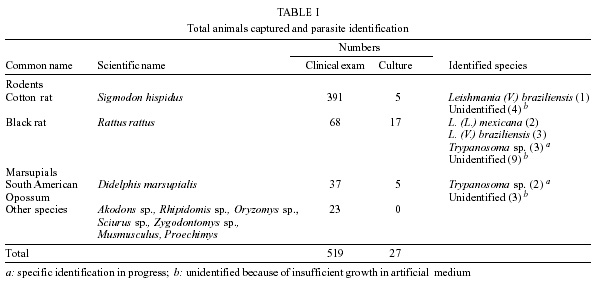
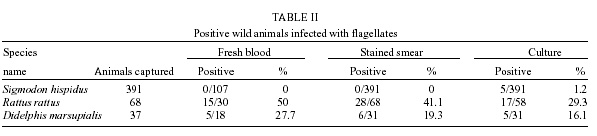
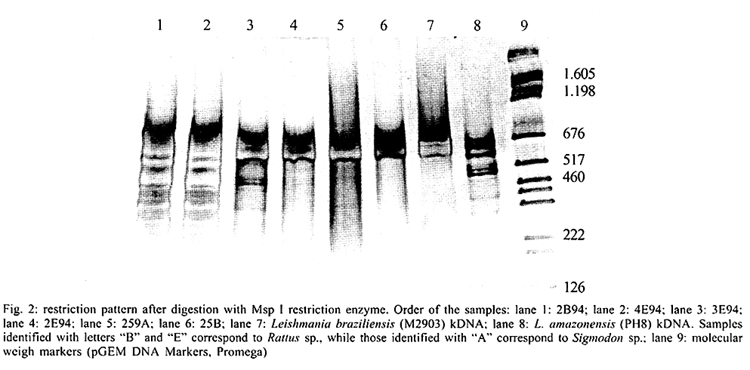
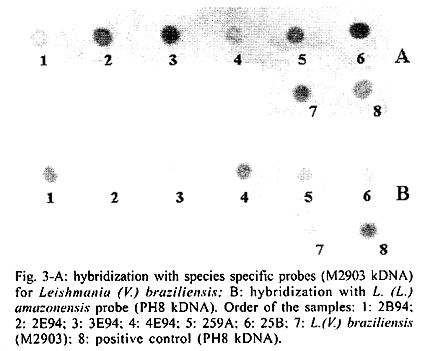
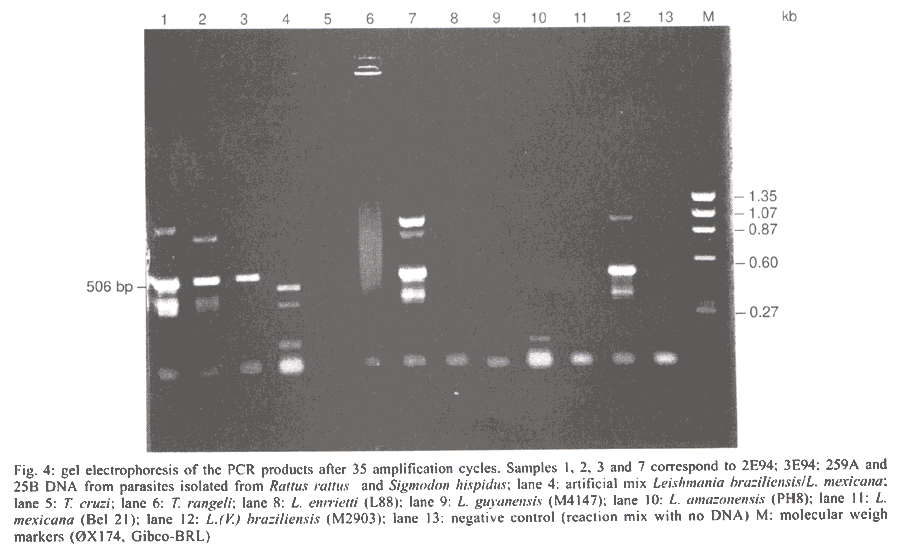
San Nicolas, San José, Apdo Postal 4043, Caracas 1010A, Venezuela Supported by the World Bank/Venezuelan government, projects Ven/96/002/016 and Ven/96/002/021-024. Received 15 May 2001 Code Number: oc02031 A total of 519 wild animals belonging to eleven species were collected during a two year study in a cutaneous leishmaniasis endemic area in Venezuela (La Matica, Lara State). The animals were captured in home-made Tomahawk-like traps baited with maize, bananas or other available local fruits, and parasites were isolated from 27 specimens. Two different species were found naturally infected with flagellates, i.e., cotton rats (Sigmodon hispidus) and black rats (Rattus rattus). Characterization of the parasites using PCR, kDNA restriction pattern and hybridization with species-specific probes revealed the presence of Leishmania (L.) mexicana in three of the black rats and Leishmania (V.) braziliensis in two others. The latter species was also identified in the single positive specimen of S. hispidus. The results suggested both species of animals as possible reservoirs of Leishmania sp. Key words: Leishmania - reservoirs - leishmaniasis - polymerase chain reaction - Venezuela Cutaneous leishmaniasis is produced by different species and subspecies of Leishmania of the subgenera Viannia and Leishmania, which infect a wide variety of mammalian hosts. In Venezuela, during the last five years, an annual total of 2,500 new cases have been reported by the Ministry of Health. Cutaneous leishmaniasis is most abundant in rural and suburban areas and is commonest in people with scarce economic resources, dedicated to agricultural activities. The disease is widely distributed in the whole country, especially in the Andean region, dealing with 60% of the total cases reported by the National Registry of Health. Although leishmaniasis has been described as a zoonosis, the natural reservoirs in most of the endemic areas are unknown because of the difficulties in isolating and identifying parasites from wild animals. Previous work carried out by Bonfante-Garrido et al. (1981) demonstrated the presence of L. braziliensis in domestic animals but with clinical manifestations similar to human disease. The relationship of this parasite with wild animals has not yet been demonstrated. Other studies mainly restricted to domestic animals and experimental infections have been reported in Venezuela (Torrealba et al. 1972, Aguilar et al. 1984). Many mammals have been described as hosts of Leishmania and some of them are considered to play an important role as reservoirs (Lainson & Shaw 1987). In Brazil, sylvatic rodents as well as edentates and marsupials are reported to be the natural reservoir hosts of dermotropic leishmaniasis (Arias et al. 1981, Lainson et al. 1981). In Venezuela, despite the high incidence of cutaneous leishmaniasis in the rural population and efforts to control the disease, there have been few epidemiological studies and in the majority of the foci the natural vectors and reservoirs are unknown. The studies in wild animals have been limited by the number of animals studied or due to the techniques used in the identification of the parasites isolated from animals. Finding the natural host of Leishmania is crucial to determine the natural cycle of the parasite, in order to understand the epidemiology of the disease. In this study we used molecular techniques to identify the parasites isolated from wild animals and incriminate wild animals as natural hosts of Leishmania spp. in an endemic area in Lara State, Venezuela. MATERIALS AND METHODS Study area - La Matica is an small village located between latitudes 9º42'50"N and longitudes 69º41'53"W in the Andres Eloy Blanco district in the southeastern sector of the Venezuelan state of Lara (Fig. 1). The district has a population of 39,842 inhabitants; the village and the surrounding areas which have an estimated population of 1,500 inhabitants and an economy based on agricultural activities, reported 10 cases of cutaneous leishmaniasis during this study. Lara State is located in west central Venezuela with a population of 1,522,043 inhabitants. It is a highly endemic area for cutaneous leishmaniasis with an incidence of 5 new cases/100,000 inhabitants/year, the state being responsable for about 20% of the new cases registered in Venezuela each year. Animal trapping - The animals were captured five nights per month over a period of two years from October 1996 to October 1998, trapped in home-made Tomahawk-like one door metal traps of two different dimensions, 19x5x5 and 26x9x9 inches, with the capacity to capture single small animals or several animals at the same time. Traps were baited with local fruits (banana, maize, etc.) set at 20 or 30 m from the houses and in the cornfields from 6 p.m. to 8 a.m. Twenty traps were used to captured an average of five animals per night; all animals were keep alive until they were studied. Diagnostic methods - The animals were examined, looking for suspicious lesions such as skin ulcers, scars, and alopecia. Blood samples were taken from the heart under anaesthetic conditions. Each sample was used for fresh microscopic examination and culture. If any of the samples resulted positive, the animal was humanely sacrificed and samples from skin, liver and spleen were taken for parasitological studies, microscopic preparation, PCR and culture in blood agar medium. Parasite isolation and culture - The parasites were isolated from blood and cultured in blood agar base medium (Difco), with 10% defibrinated rabbit blood plus 200 units of penicillin. Within seven days, samples of the cultures were taken for microscopic examination. Negative cultures without contamination were kept for one month before discarding. When the culture was positive, rapid growth of the parasites was obtained in petri dishes in the same medium. Parasites in the stationary phase were harvested by centrifugation at 5,000 rpm for 10 min at 4oC and washed three times with sterile phosphate buffered saline (pH 7.4) KDNA isolation and digestion - Kinetoplast DNA was isolated as previously described (Rodriguez et al. 1994). For schizodeme analysis, 5 µg of kDNA were digested with five units of Msp I endonuclease (Gibco-BRL). After digestion to completion, the digested products were separated by electrophoresis in 4.5-10% linear gradient polyacrylamide gels, run at 7 mAmp overnight. The gels were stained with silver nitrate and photographed. Dot blot hybridization was carried out as was previously described (Barrios et al. 1994). Pre-hybridization and hybridization were performed overnight at 42oC in 50% formamide containing buffer. Filters were developed using Genius Systems (Boehringer-Mannheim) according to the instructions of the manufacturer. PCR reaction - For the PCR, DNA was extracted from biological samples after digestion in lysis buffer (10mM Tris-HCl pH 8, 10mM EDTA, 1 µg/ml Proteinase K) incubated for 2 h at 56oC followed by phenol/chloroform extraction and ethanol precipitation. The DNA was precipitated from the aqueous phase and resuspended in TE buffer (10 mM Tris HCl, pH 8, 1 mM EDTA). PCR was carried out with 5 µl of total DNA using the same conditions and primers previously described (Rodriguez et al. 1997). RESULTS Live trapping - The species and number of wild animals trapped are shown in Table I. Animals were captured during five days periods each month for two years and identified using taxonomic criteria. Most of the animals captured were rodents (482) or opossums (37), cotton rats (S. hispidus) accounted for 391 animals followed by black rats (R. rattus) with 68 animals. Others species found were Akodon sp. (10), Rhipidomis sp. (2), Oryzomys sp. (3), Sciurus sp. (3), Zygodontomys sp. (1), Mus musculus (1) and Proechimys sp. (3). Opossums were all Didelphis marsupialis (37/37). Clinical and parasitological diagnosis - The results obtained using different diagnostic criteria are shown in Table II. Comparison between microscopic observation and culture indicated that culturing of the parasites was more sensitive than smear and microscopic observation after Giemsa staining in the detection of infected S. hispidus. Microscopic examination of fresh blood was more sensitive than other methods in the identification of parasites in blood samples from Didelphis sp. and R. rattus, but was not able to detect infected S. hispidus. Smears from lymph node, liver, spleen and skin were negative for amastigotes. Blood cultures were successfully used to obtain 27 isolates (Table II), from three different species of mammals; S. hispidus (5/391), R. rattus (17/58) and Didelphis sp. (5/31). Only eleven cultures grew in artificial culture medium, to be characterized using molecular techniques. Taxonomic identification of parasites - The restriction patterns of six isolates after digestion with Msp I restriction enzyme are shown in Fig. 2. Samples 1 and 2 have similar restriction patterns with L. (L.) amazonensis (lane 8, IFLA/BR/68/PH8), while isolates in lanes 3, 4, 5 and 6, have high homology with L. (V.) braziliensis (MHOM/BR/68/M2903) in lane 7. These results were confirmed after hybridization with probes for L.(V.) braziliensis and L.(L.) mexicana respectively. The results after hybridization with L.(V.) braziliensis probe (M2903 kDNA) are shown in Fig. 3A. A high hybridization signal was observed in samples 2, 3, 5 and 6, corresponding to the same samples in Fig. 2. In Fig. 3B the high hybridization signals correspond to 2B94 and 4E94 (samples 1 and 4), after hybridization with a L. (L.) amazonensis (PH8) kDNA probe. Both samples were isolated from black rats. PCR results - The results obtained after 35 amplification cycles are shown in Fig. 4. The expected diagnostic band of 506 bp for L. (V.) braziliensis was observed in three samples from black rats (lanes 1, 2 and 3) and one sample from S. hispidus (lane 7). Two other samples from black rats (4, 5) resulted negative for L. braziliensis, but PCR with L. mexicana primers was positive for those samples (result not shown). Although the diagnostic band was observed with a high intensity signal, other bands were also observed, probably due to genetic variability within the L. (V.) braziliensis species as we previously described (Rodriguez et al. 1998). Hybridization of the positive PCR products to LbJ38 L. braziliensis species-specific probe resulted positive (results not shown). DISCUSSION Many species of wild animals have been suggested as reservoirs of New World Leishmania spp. (Lainson & Shaw 1987). Amastigotes of Leishmania have been isolated from mammals of many species and several orders. The reservoirs for L. (V.) guyanensis include the two-toed sloth, Choloepus didactylus and the lesser anteater, Tamandua tetradactyla; secondary hosts include marsupials (Didelphis) and more rarely rodents (Proechimys) (Lainson et al. 1994), while L.(V.) braziliensis has been isolated from the rodents Proechimys, Rhipidomys, Oryzomys, Akodon and Rattus as well as Didelphis. In Venezuela, an early report suggested Zymodontomys and Heteromys to be the reservoirs for L. pifanoi, based on experimental infections in hamsters after injection with blood isolated from both animals (Kerdel-Vegas & Essenfield-Yahr 1966). Domestic animals such as equines and cats have been reported infected with L. (V.) braziliensis (Bonfante et al. 1981). Here we report the results obtained after a field study carried out in an endemic area of cutaneous leishmaniasis in Lara State, Venezuela. A total of 519 mammals was trapped including 11 different species. The most abundant were cotton rats (S. hispidus) accounting for 391 specimens and black rats (R. rattus), with 68 specimens. However, the presence of parasites was only demonstrated in 27 mammals after culturing blood samples from the most frequently captured species. The kDNA restriction pattern together with the hybridization results indicate that four samples were infected with L. (V.) braziliensis (one cotton rat and three black rats). L. (L.) mexicana was demonstrated in two samples from black rats. The PCR and hybridization results using species-specific primers and probes also confirm the presence of both of the Leishmania and Viannia subgenera in wild animals. The utility of the mentioned molecular tools in the specific identification of L. (V.) braziliensis was previously demonstrated (Rodriguez et al. 1997). kDNA restriction pattern and hybridization are techniques widely used to differentiate between species and sub-species of parasites (Barker 1989, Rodriguez et al. 1994, Pacheco et al. 1997). Here we demonstrated once again the utility of both techniques to identify Leishmania in samples isolated from wild mammals. Black rats (R. rattus) were previously identified as the natural reservoir for Leishmania in the State of Ceará, Brazil (Alencar et al. 1960, Vasconselos et al. 1987). More recently the same species was identified as a natural reservoir for L. (V.) braziliensis in Baturite, Brazil (Vasconselos et al. 1994) and in Colombia (Alexander et al. 1998), using PCR and hybridization techniques. In this study, the results suggest that cotton rats and black rats are putative reservoirs of L. (V.) braziliensis and L. (L.) mexicana. Both species meet at least three of the five criteria used to define a reservoir: (1) overlap between the geographic distribution of wild animals and human cases. Cutaneous leishmaniasis was reported in the study area and the parasites isolated from the human cases were identified as L. (V.) braziliensis (Rodriguez et al. 1997); (2) proportion of wild animals infected: it may be higher than 20%. In this study we found that 17/68 balck rats were infected, representing 25% of the specimens; (3) the identification of L. braziliensis in both species of animals. Additionally, xenodiagnosis using Lu.ovallesi reveals 1/500 sandflies with parasites in the digestive tract after biting an infected S. hispidus. Chable Santos et al. (1995) demonstrated the role of cotton rats in the transmission of L. mexicana in Mexico; our findings suggest the same animal as a possible reservoir of Leishmania spp. The results presented here offer additional evidence about the role of black rats in the transmission and maintenance of cutaneous leishmaniasis as a zoonosis. We believe that this is the first demonstration of both species of mammals as natural hosts of Leishmania of both of the subgenera Viannia and Leishmania in Venezuela. Finding the same species (R. rattus) infected with L.(L.) mexicana or L.(V.) braziliensis is very interesting from the epidemiological point of view, since it has been previously demonstrated (Barrios et al. 1994) that the same sandfly can be the vector for both subgenera. It is also possible that the same animal is bitten by two different species of vector. However, more studies have to be done to confirm the role of both species as natural reservoirs of Leishmania spp. in Venezuela. ACKNOWLEDGEMENTS To Mr Felix Flores and Wilmer Galindo for technical assistance and to the community of La Matica for support given during field work, also to Dr Marian Ulrich for the corrections and critical reading of the manuscript. REFERENCES
© 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02031f4.gif] [oc02031f2.jpg] [oc02031f4.jpg] [oc02031t2.jpg] [oc02031t1.jpg] [oc02031f3.jpg] [oc02031f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}