 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(2) 2002, pp. 221-225 Stenurus globicephalae Baylis et Daubney, 1925 (Nematoda: Pseudaliidae) from a False Killer Whale, Pseudorca crassidens (Cetacea: Delphinidae), Stranded on the Coast of Uruguay María Inés Zylber/+, Gabriela Failla*, Alfredo Le Bas** Laboratorio de Helmintología,
Departamento de Ciencias Biológicas, Facultad de Ciencias Exactas y Naturales,
Universidad de Buenos Aires, Ciudad Universitaria, Pabellón II, 4º
piso, (C1428EHA) Buenos Aires, Argentina *Laboratorio de Zoología de
Invertebrados **Sección Fisiología, Departamento de Biología
Animal, Facultad de Ciencias, Universidad de la República, Montevideo,
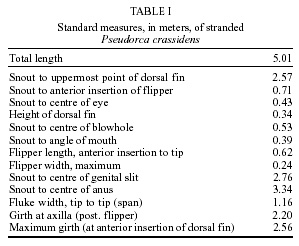
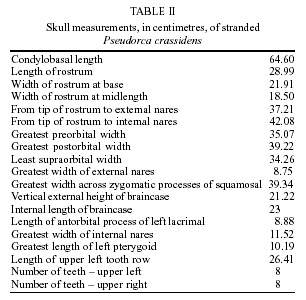
Uruguay Financial support: ANPCyT (Project BID 1201 OC-AR PICT 1-6604) and CONICET (Project PIP 4545/96) Received 12 June 2001 Code Number: oc02042 Stenurus globicephalae Baylis et Daubney, 1925 (Nematoda: Pseudaliidae) was found in the cranial air sinuses of a false killer whale, Pseudorca crassidens (Owen), stranded on the coast of Uruguay in 1999. Although this species has been reported once in P. crassidens from the North Atlantic, this is the first record for South America. A total of 920 specimens were obtained, of which 663 were females (body length: 4.34 ± 0.45 cm) and 257 were males (2.99 ± 0.18 cm). Morphometric details are presented for S. globicephalae in this host, which do not show significant differences from those parasitizing Globicephala melas (Traill), but are distinct from those parasitizing Peponocephala electra (Gray). The host's skull revealed loss of osseous mass with the disappearance of the left zygomatic arch, and the left jaw had three osseous fenestrations in the region related to the organ of acoustic reception. These lesions support the hypothesis that this infection, known as stenurosis, was related to the stranding. Key words: Pseudorca crassidens - stranding - Pseudaliidae - Stenurus globicephalae - taxonomy - Uruguay The false killer whale, Pseudorca crassidens (Owen) is a gregarious delphinid and often forms schools containing more than a hundred individuals. It is distributed in deep offshore tropical to warm temperate waters and generally does not range further north or south than 50º in either hemisphere (Jefferson et al. 1993). This is one of the most common species implicated in cetacean mass strandings, with schools of up to 835 individuals involved (Nowak 1991). There are many suggestions as to the cause of mass strandings. Some authors mention the strong social cohesion of a pod as being the reason why healthy animals may follow a sick individual on to the beach (Rogan et al. 1997). On March 10-14, 1999, a massive stranding of approximately 40 individuals of false killer whales occurred on the coast at Jaureguiberry (34º53'84''S; 55º15'33''W), Uruguay. Thirty of the stranded animals were returned alive to the ocean, while dead ones were quickly buried by local authorities and non governmental organizations. On March 13, 1999, an adult male was found dead on Punta Colorada beach, 18.9 km from this massive stranding, and was examined for pathology related to its death. The sole anomalies detected were the presence of abundant lungworms (pseudaliid nematodes), identified as Stenurus globicephalae Baylis et Daubney, 1925, in cranial air sinuses, and of osseous alterations in the left jaw and the left zygomatic arch. Most species of Stenurus are found in the middle ear, eustachian tube and cranial sinuses, while a few of them are found in bronchi and bronchioles (Measures 2001). Species of Stenurus have been reported in various odontocete hosts throughout the world, including the false killer whale. Odell et al. (1980) found S. globicephalae in the pterygoid sinus complex of five and in the lungs of four animals stranded in Florida, U.S.A. Research on parasites of South American marine mammals is scarce and there are no records of the helminth fauna of P. crassidens from this region. In the current study, we report for the first time the presence of S. globicephalae in a delphinid from South America and add a short description and new morphometric characters for this parasite in P. crassidens. Additionally, its possible relation to the stranding is discussed. MATERIALS AND METHODS A partial necropsy of the false killer whale was carried out in the place of stranding. After taking morphometric data, the abdominal cavity was examined in order to search for any pathology. The thorax was not opened, therefore lungs and heart could not be inspected. The cranium was disarticulated from the trunk at the atlas-occipital condyles. Removal of soft parts allowed air sinuses to be examined. All available nematodes found on the mucous coating of both left and right sinuses were collected, fixed in 10% isotonic formaline solution and preserved in 70% ethanol. Specimens were cleared with Amann's lactophenol or beechwood creosote in temporary mounts for morphological study. En face views were mounted and studied in lactophenol. Worms were examined morphologically and 20 specimens of each sex (n) measured using a Carl Zeiss Axioskope microscope equipped with a drawing tube. All measurements are given in micrometers (µm), unless otherwise stated, with the range followed by the mean in parenthesis. Voucher specimens, BP/9002 (females) and BP/9003 (males), were deposited in the Helminthological collection, Laboratorio de Zoología de Invertebrados, Facultad de Ciencias, Montevideo, Uruguay. After maceration of soft parts, the skull was inspected for bone lesions, measured and deposited in the collection of the Sección Zoología de Vertebrados, Facultad de Ciencias, Montevideo, Uruguay (ZVM 1813). RESULTS Parasites were identified as S. globicephalae after Anderson (1978), Baylis and Daubney (1925), and Arnold and Gaskin (1975). Out of the 920 specimens collected, 257 were males and 663 were females, corresponding to a sex ratio of 1: 2.58. There is a marked sexual dimorphism: females are longer (4.34 ± 0.45 cm) and more stout than males (2.99 ± 0.18 cm), and present a somewhat curved posterior extremity. Metastrongyloidea Pseudaliidae Railliet & Henry, 1909 Stenurinae Skrjabin, 1942 Stenurus Dujardin, 1845 Stenurus globicephalae Baylis
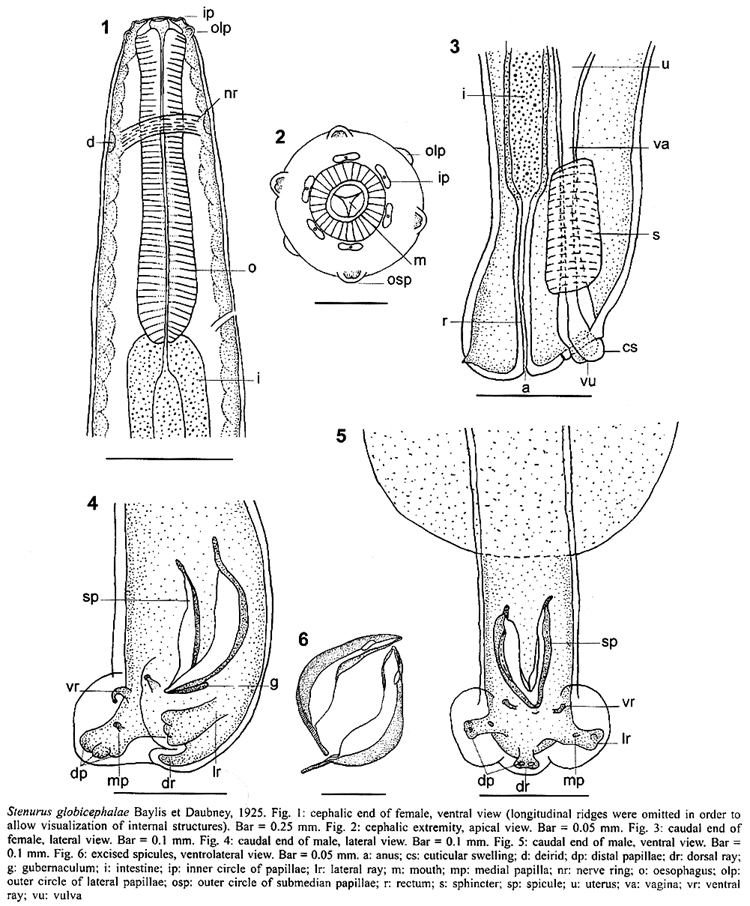
et Daubney, 1925 Redescription: body stout in its anterior extremity, tapering posteriorly. Buccal cavity funnel-shaped, with thick cuticularized walls (Fig. 1). Mouth surrounded by inner circle consisting of six papillae and an outer circle consisting of two lateral and four submedian papillae (Fig. 2). Oesophagus club-shaped in its posterior part. Longitudinal cuticular ridges arise posteriorly to the cephalic region. Female (Fig. 3): body 38.9-47.9 (43.4; n = 60) mm long, 560-660 (616) maximum width. Buccal capsule 40-65 (45) wide in anterior and 75-90 (80) in posterior part. Diameter of cephalic end at level of posterior cephalic papillae 115-150 (132). Oral capsule + oesophagus 520-700 (610) long, 95-130 (101) maximum width. Nerve ring 170-250 (198), deirids 200-290 (232), and excretory pore 490-650 (539) from anterior end, respectively. Posterior end truncated, with subterminal protuberances. Vulva 40-75 (56) from anus, with spherical cuticular swelling on anterior lip in some specimens. Anus 40-75 (53) from posterior end. Vaginal sphincter elongated, 90-110 (98) long, 45-55 (50) wide. Eggs 37.5-55 (49; n = 50) long, 25-45 (32; n = 50) wide containing first stage larvae in genital ducts. Male (Figs 4, 5, 6): body 28.1-31.7 (29.9, n = 60) mm long, 320-420 (384) maximum width. Buccal capsule 25-40 (32) wide in anterior and 55-75 (67) in posterior part. Diameter of cephalic end at level of posterior cephalic papillae 105-135 (117). Oral capsule + oesophagus 500-600 (536) long, 80-100 (91) maximum width. Nerve ring 160-200 (172), deirids 190-250 (207), and excretory pore 475-550 (516) from anterior end, respectively. Cuticle considerably swollen in front of bursa copulatrix, forming two prominent, globular alae. Bursa not distinctly lobed, consisting of five rays distributed in two small, thin ventral rays 15-25 (19) long; two stout, trifid lateral rays 32.5-50 (40) long, with a stalked medial ventral and three sessile distal papillae, and one short, broad dorsal ray 20-35 (24) long, with two ventrolateral papillae at tip. Spicules equal in size, short and curved, 110-135 (116) long. Gubernaculum 40-65 (50) long Host: Pseudorca crassidens (Owen) (Cetacea: Delphinidae) Site of infection: cranial air sinuses Locality: Jaureguiberry Beach (34º53'84''S; 55º15'33''W), Uruguay Specimens deposited: BP/9002 (females) and BP/9003 (males) (vouchers) The host was an adult male 5.01 m long. Its morphometric data are presented in Table I. No anomalies were found in the abdominal cavity. The host's skull revealed loss of osseous mass with the disappearance of the left zygomatic arch, while the tympanic bones showed no anomalies. The left jaw had three osseous windows (81.6 x 11.1; 77.7 x 8.2 and 15.3 x 1.9 mm) in the region related to the organ of acoustic reception. Skull measurements are displayed in Table II. DISCUSSION At present, eight species are considered as valid members of the genus Stenurus: S. minor (Kühn, 1829) Baylis et Daubney, 1925; S. globicephalae Baylis et Daubney, 1925; S. ovatus (Linstow, 1910) Baylis et Daubney, 1925; S. arctomarinus Delyamure et Kleinenberg, 1958; S. truei Machida, 1974; S. nanjingensis Tao, 1983; S. yamaguti Kuramochi, Araki et Machida, 1990, and S. australis Tantalean et Sarmiento, 1991. A comparative study showed that morphological characteristics of the nematodes obtained from P. crassidens correspond to S. globicephalae, first described from pilot whales [Globicephala melas (Traill)] in the North Atlantic and redescribed by Kikuchi and Nakajima (1996) from melon-headed whales [Peponocephala electra (Gray)] in Japan. It has also been reported from the same and several other delphinids such as Globicephala macrorhynchus Gray, Lagenorhynchus acutus (Gray), Grampus griseus (Cuvier), Feresa attenuata Gray and P. crassidens from NW and NE Atlantic, SW and SE Pacific and the Caribbean Sea (Cowan 1967, Dailey & Brownell 1972, Arnold & Gaskin 1975, Cannon 1977, Beverley-Burton 1978, Forrester et al. 1980, Odell et al. 1980, McManus et al. 1984, Raga & Balbuena 1987, Morales-Vela & Olivera-Gómez 1993, Rogan et al. 1997, Abollo et al. 1998, Gibson et al. 1998, Mignucci-Giannoni et al. 1998, Measures 2001). This is the first report of S. globicephalae in South American waters and the first of the genus Stenurus from the South Atlantic. From South America only S. australis has been previously reported, on the Pacific coasts of Peru and Chile, in the porpoise Phocoena spinipinnis (Burmeister, 1865) (Torres et al. 1994). The present study presents the morphometric characters of specimens found in a different host and geographical region than previous descriptions. The original description by Baylis and Daubney (1925) of S. globicephalae does not discriminate between sexes in some measurements and lacks many others. There are not significant morphometric differences between S. globicephalae parasitizing G. melas and these parasitizing P. crassidens, except for the absence of the cuticular appendage (probably broken due to manipulation) on the anterior lip of the vulva of the majority of our female specimens. Nematodes, both males and females, parasitizing P. electra are smaller in total body length and maximum body width, although the values of the rest of the measurements are larger or equal. Pseudaliids are known to cause granulomatous inflammation in the lungs of cetaceans (verminous bronchopneumonia), a common cause of death (Cowan 1967, Cannon 1977, Dailey & Stroud 1978, Odell et al. 1980). Some adult pulmonary pseudaliids, including S. globicephalae, stimulate none or little inflammatory response in bronchi or bronchioles, but larvae in alveoli can cause a subacute purulent focal pneumonia (Cowan 1967, Dailey & Stroud 1978, Measures 2001). Pseudaliids located in the cranial sinuses and middle ear provoke minor hemorrhage, mild to moderate nonsuppurative chronic inflammation and thickening of the sinus mucosal lining, rarely purulent sinusitis (Delyamure 1955, Measures 2001). The disease caused by Stenurus spp. is known as stenurosis and it is distributed worldwide among dolphins. The presence of these lungworms, often packed together in huge numbers, has been associated with obstruction of the auditive ducts and with osseous lesions, leading to the hypothesis that this infection could represent a potential factor in the stranding of Odontoceti (Delyamure 1955, Dailey & Stroud 1978, Dailey & Walker 1978, Morimitsu et al. 1992). According to Parry et al. (1983), specimens of S. globicephalae in pilot whales occur in the head sinuses and middle ear, but not in the inner ear where the ultrasonic echoes of their sonar are detected. However, large numbers of moving nematodes in the cranial sinuses, close to the inner ear, may affect hearing, interfere with the echolocative function or at least result in a loss of concentration, bringing about subsequent negative effects on diving performance, ability to detect fishing nets, food catching capability, navigation and body condition (Morimitsu et al. 1987, Gibson et al. 1998, Faulkner et al. 1998, Measures 2001). The possible pathogenicity of S. globicephalae in cetacean hosts at present is not clearly understood. The osseous lesions in aerial cavities related to the middle ear and in the jaw of the specimen examined suggest important alterations in the organ of acoustic reception and, thus, support the hypothesis that the disorientation which caused the stranding in this case could be related to stenurosis. ACKNOWLEDGEMENTS To "SOS Rescate de Fauna Marina", especially to its Director, Mr Richard Tesore, for allowing us to use the installations of the rescue centre for the fieldwork. To Lic. Alicia Gil for critical revision of this manuscript and to the cetologist Ricardo Praderi and to Dr Lena Measures for facilitating specific bibliography. REFERENCES
© 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02042f1-6.jpg] [oc02042t2.jpg] [oc02042t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}