
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 2, 2002, pp. 281-283
|
Memórias do Instituto Oswaldo Cruz -
Mem
Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(2) 2002, pp. 281-283
SHORT COMMUNICATION
Thermal
Constant of an Experimental Population of Muscina stabulans (Fallén 1817)
(Diptera:Muscidae) in the Laboratory
Luciene Maura Mascarini/+,
Ângelo Pires do Prado*
Departamento de Parasitologia, Instituto
de Biociências, Unesp, Rubião Junior s/n°, 18618-000 Botucatu, SP, Brasil *Departamento
de Parasitologia, Instituto de Biologia, Unicamp, Campinas, SP, Brasil
+Corresponding author.
Fax: +55-14-6821.3744. E-mail: luciene@ibb.unesp.br
Finnancial support: Capes
Received 9 March 2001
Accepted 5 November 2001
Code Number: oc02053
A study was conducted to investigate
the thermal constant in an experimental population of Muscina stabulans
(Fallén 1817). A natural population was obtained from a livestock and their
F1 generations were maintained at four constant temperatures (16°C,
20°C, 26°C and 31°C). The thermal constant was calculated by the hyperbole method
using the base temperature, tb = 4.4°C and the thermal constant K = 35.3 GD.
Ricker's geometric regression, considering tb = 7.8°C and K = 28.9 GD was also
calculated.
Key words: Muscina stabulans -
thermal constant - constant temperature
Muscina stabulans (Fallén
1817) (Diptera: Muscidae) a common eusinanthropic fly in farms and orchards,
may be associated with human dwellings (Smith 1986). In addition, it can play
a role in the control of some Diptera species because of its predatory habits
on their larvae (Legner & Dietrich 1989). The growth pattern and development
of insects are influenced by temperature and can be measured under laboratory
conditions. It is possible to estimate the number of generations per year and
the potential ability of population increase of at a specific temperature by
measuring developmental period, mortality and oviposition pattern under constant
temperature conditions (Howe 1967). The thermal requirements of an insect can
be appraised by the thermal constant (K), expressed in degree-days (Haddad &
Parra 1984). This method has been utilized for many years in forecasting the
occurrence of infestations in agriculture and the thermal constant can be used
as indicators of the distribution and abundance of insect populations. The objective
of this study was to establish a thermal constant for a population of M.
stabulans.
Experimental populations of M.
stabulans were obtained from livestock farm Capuavinha (26º57'S, 47º19'W),
town of Monte-Mor, SP, Brazil. These flies were transferred to cages (30 cm
x 15 cm x 15 cm), where they were fed yeast suggar and water and maintened at
27 ± 30°C. Two thousand eggs were collected and dividided into 40 pots
(9.5 cm x 7.5 cm) with 50 eggs/pots and 10 pots (replicates) at each temperature:
16°C, 20°C, 26°C and 31°C, tainning 200 g of fermented of Labina-Purina mouse
ration. To these pots, a further 300 g of mouse ration was added and the larvae
allowed to grow until pupation. Pupae were removed and transferred to other
cages, where they hatched. The total development time was recorded. The basic
development temperature to calculate the thermal constant was determined by
the hyperbole method (Bean 1961), using the total development times from the
four constant temperature groups. Thermal constant was obtained from the equation
K = D(T-Tb), where K = constant thermal, D = development time (hours), T = temperature
in which the insect grew and Tb = basic temperature of the insect, obtained
from the Healthy linear regression program (1986) and from Ricker's geometric
average (1986) regression program (Krebs 1989).
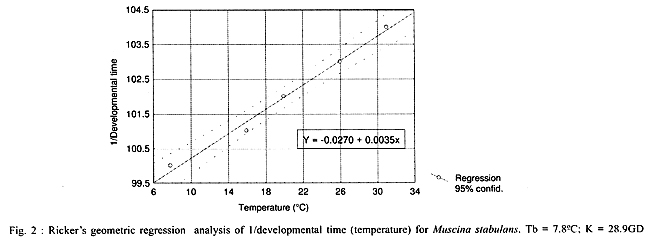
The Table
shows the total development time from larval to adult stage. Average larval
development times decreased with increasing temperature. This was inversely
proportional to increase in temperature, except at 31°C when development time
was longer than at 26°C. The data obtained at 26ºC are similar to those obtained
by Kobayashi and Mizushina (1937) with 14.06 days of total development time,
Vinogradova and Marcenko (1984), 12.9 days, and Krzywinski (1993) who analyzed
the thermal constant at 28ºC, obtaining an average of 12.9 to 14 days. The total
development time, observed by Lewallén (1952) at 26.7ºC, was longer (20-25)
that obtained in our study (12.8 days). Thermal requirements were estimated
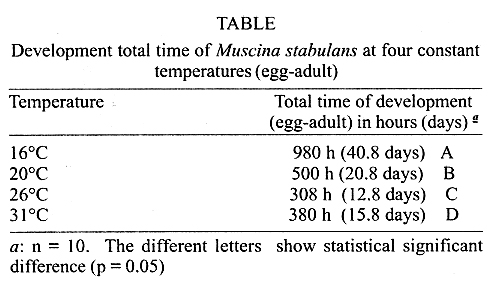
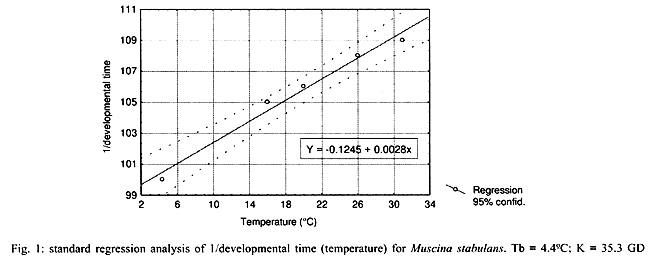
by determining the thermal constant K and expressed in degree-days. Fig. 1 shows the regression analysis,
which showed Tb = 4.4°C and K = 35.3 GD, and Fig. 2 shows Ricker's regression with
Tb = 7.8°C and K = 28.9 GD.
Insect body temperature is probably
more important than environmental temperature in influencing behavior (Chapmam
1983). Temperature limits can be established by inactivity and by the destruction
of some enzymes (Howe 1967). Lethal temperature limits are metabolically regulated
by enzymes. Metabolic pattern is measured by oxygen consumption, and when ambient
temperature is close to the maximum limit, metabolism is reduced, probably due
to denaturation of enzymes (Chapman 1983). The knowledge of the thermal requirements
of insects estimated by basic temperature and thermal constant permits, temporarily,
store insects under certain artificial conditions, facilitating study program
and field work (Aguiar-Valgorde & Milward Azevedo 1992). The threshold and
thermal constant can also be used as indicators of distribution and abundance
of insect populations (Messenger 1959). From these models, temperature occupies
a prominent place, because it is a metodologic component that affects insect
populations and is easy to predict.
REFERENCES
- Aguiar-Valgorde M, Milward-Azevedo
EMV 1992. Determination of the thermal demands of Stomoxys calcitrans
(L.) (Diptera:Muscidae), in laboratory conditions. Mem Inst Oswaldo Cruz
87: 11-20.
- Bean JL 1961. Predicting emergence
of second instar budworm larval from hibernation under field conditions in
Minesota. Ann Entomol Soc Am 54: 175-177.
- Chapman RF 1983. The Insects:
Structure and Function, Hodder & Stoughton, London, 919 pp.
- Hadad ML , Parra JPR 1986. Métodos
para Estimar os Limites Térmicos e a Faixa Ótima de Desenvolvimento nas Diferentes
Fases do Ciclo Evolutivo dos Insetos, Agricultura e Desenvolvimento, ESALQ,
EDUSP, São Paulo, 12 pp.
- Healthy Insittute Inc 1986. Healthy
User's Guide: Statistics, 6th ed., Cary, North Carolina, 200 pp.
- Howe RW 1967. Temperature effect
on embrionic development in insects. Ann Ent Rev 12: 15-41.
- Kobayashi H, Mizushima H 1937.
The relationship between the laboratory temperature and development of flies.
K J Med 8: 19-39.
- Krebs J 1989. Ecological Methods,
Harper & Row, NY, 654 pp.
- Krzywinski J 1993. Development
of two Muscina species (Diptera, Muscidae) at constant temperature.
Bull Entomol Pologne 62: 125-132.
- Legner EF, Dietrick EJ 1989. Coexistence
of predatory Muscina stabulans and Ophyra aenescens (Diptera:
Muscidae) with dipterous prey in poultry manure. Entomophaga 34:
453-461.
- Lewallen LL 1952. Laboratory studies
of the false stable fly. J Econ Entomol 45: 515-517.
- Messenger PS 1959. Bioclimatic
studies with insects. Ann Rev Ent 4: 183-206.
- Ricker WE 1984. Computation and
use of central trend lines. Can J Zool 62: 1897-1905.
- Smith KGV 1986. A Manual of
Forensic Entomology, British Museum and Cornell University Press, London,
205 pp.
- Vinogradova E, Marcenko VM 1984.
Ispol'zovanie temperaturnych parametrov razvitija much v sudebno-medicinskoj
praktike. Sud-Med Ekspert 1: 16-19.
© 2002
Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02053f2.jpg]
[oc02053f1.jpg]
[oc02053t1.jpg]
|

{kind=link}
{kind=link}
{kind=link}