 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(3) 2002, pp. 307-311 Ultrastructural, Antigenic and Physicochemical Characterization of the Mojuí dos Campos (Bunyavirus) Isolated from Bat in the Brazilian Amazon Region Ana LM Wanzeller/+, José AP Diniz, Maria LC Gomes, Ana CR Cruz, Manoel CP Soares, Wanderley de Souza*, Amélia PA Travassos da Rosa**, Pedro FC Vasconcelos Instituto Evandro Chagas, Av. Almirante
Barroso 492, 66090-000 Belém, PA, Brasil *Laboratório de Ultraestrutura
Celular, Instituto de Biofísica Carlos Chagas Filho, Universidade Federal
do Rio de Janeiro, Rio de Janeiro, RJ, Brasil **Department of Pathology, University
of Texas Medical Branch, Galveston, TX, USA This work is part of the senior author's MSc thesis and was accomplished with the financial support of Conselho Nacional de Desenvolvimento Científico e Tecnológico, Capes, Universidade Federal do Rio de Janeiro, Universidade Federal do Pará and Instituto Evandro Chagas/Fundação Nacional de Saúde. Receivd 23 April 2001 Code Number: oc02059
The Mojuí dos Campos virus (MDCV) was isolated from the blood of an unidentified bat (Chiroptera) captured in Mojuí dos Campos, Santarém, State of Pará, Brazil, in 1975 and considerated to be antigenically different from other 102 arboviruses belonging to several antigenic groups isolated in the Amazon region or another region by complement fixation tests. The objective of this work was to develop a morphologic, an antigenic and physicochemical characterization of this virus. MDCV produces cytopathic effect in Vero cells, 24 h post-infection (p.i), and the degree of cellular destruction increases after a few hours. Negative staining electron microscopy of the supernatant of Vero cell cultures showed the presence of coated viral particles with a diameter of around 98 nm. Ultrathin sections of Vero cells, and brain and liver of newborn mice infected with MDCV showed an assembly of the viral particles into the Golgi vesicles. The synthesis kinetics of the proteins for MDCV were similar to that observed for other bunyaviruses, and viral proteins could be detected as early as 6 h p.i. Our results reinforce the original studies which had classified MDCV in the family Bunyaviridae, genus Bunyavirus as an ungrouped virus, and it may represent the prototype of a new serogroup.
Key words: arbovirus - Mojuí dos Campos virus - characterization - ultrastructure - protein kinetics - Amazon region - Brazil Arboviruses form an ecological group of viruses transmitted by arthropods to man and other animals. Some types are associated with important public health problems at regional, national and/or continental levels (Vasconcelos et al. 1992). In Brazil, at least 200 distinct serotypes have been isolated. Of these, 186 were obtained from the Amazon region of Brazil, and 86 of them have been considered as new types (Travassos da Rosa et al. 1998). The Bunyaviridae family comprises over 300 individual virus species belonging to five genera: Bunyavirus, Hantavirus, Nairovirus, Phlebovirus and Tospovirus (Gonzalez-Scarano & Nathanson 1996). The genome of bunyaviruses consists of three segments of single-stranded, negative sense RNA that form an helical nucleocapsid by combining with the nucleoprotein. The members of the Bunyaviridae have no matrix protein and their nucleocapsid apparently interacts directly with the envelope glycoproteins, G1 and G2, during virus replication. Maturation of viral particles occurs upon Golgi or endoplasmic reticulum membranes. Exceptionally, the assembly also occurs on the plasma membrane (Madoff & Lenard 1982, Kuismanen et al. 1985, Pobjecky et al. 1986, Anderson & Smith 1987, Goldsmith et al. 1995). Mojuí dos Campos virus (MDCV) is registered in the Internacional Catalogue of Arbovirus (Karabatsos 1985) as an ungrouped, unclassified, possible arbovirus. This status was based on serologic studies carried out at Evandro Chagas Institute (Karabatsos 1985, Pinheiro & Travassos da Rosa 1985). Zeller et al. (1989) showed that MDCV had cross reaction by IFI with specific immune serum of Nyando virus (Nyando serogroup) and San Angelo (California serogroup) of Bunyaviridae family. These reactions were later confirmed by complement fixation (CF) test, but no reaction was observed by neutralization (N) tests with other viruses of Nyando and California groups (Eretmapodites 146 and Jamestown Canyon, La Crosse and Snowshoe hare). In order to establish a definitive classification to MDCV we carried out several laboratory examinations. This paper describes the results of new serological tests, culture in Vero cells, and ultrastructural features of MDCV as well as the kinetic and electrophoretic analysis of the viral proteins. MATERIALS AND METHODS
Virus isolation - MDCV was isolated from the blood of a bat, by the intracerebral inoculation of suckling mice. After four passages in these mice, the suspension of brain tissue was inoculated into Vero cells, from which an isolation was obtained. Till now, no other strain of this virus has been isolated. Virus infection - In order to infect cell cultures with MDCV, 0.1 ml of a 1:10 dilution of stock virus (about 10-7.0 TCD50/0.1 ml) was inoculated into the center of a confluent (70% to 80%) monolayer of Vero cells in 25 cm2 plastic flasks. The mixture was incubated for 1 h to allow adsorption of the virus on the cells. Cell maintenance medium (MEM; Sigma) supplemented with 2% fetal calf serum, L-glutamyne 200 mM, penicillin (100 UI/ml), streptomycin (100 µg/ml), HEPES 1M (pH 7.3), amphotericin B (3 µg/ml) and 1.5 g/l sodium bicarbonate were then added and the flasks incubated at 37°C. The cells were examined daily until the cytopathic effect (CPE) become evident. When cells showed 80% of CPE, the supernatant was collected, frozen at -70°C for 15 min, and then centrifuged at low speed (900 x g for 30 min), aliquoted and stored at -70°C with 20% fetal calf serum. Suckling mice were inoculated intracerebrally with 10-5.3 LD50/0.02 ml. When showing signals of illness they were sacrificed and the brains were immediately processed or frozen at -70°C. Preparation of antisera - Hyperimmune specific antiserum was produced in young (4-6 weeks) Swiss mice and the immunogen used was 0.2 ml of 1:10 suspension of MDCV in 0.85% NaCl solution. Four intraperitoneal injections at weekly intervals were used for immunization (Casals 1967). A week after the last immunization, the animals were anesthetized and the blood collected by intracardiac exanguination. Serum was obtained by centrifugation (2,400 g/10 min), and stored at -20° C until used. Virus titration and serology - MDCV titrations were carried out with, and without, 1:500 DCA (sodium desoxycholate) solution, and the material inoculated into newborne mice (Theiler 1957). N test in mice, IFI assay and CF test were performed using MDCV, as previously described elsewhere (Fulton & Dumbell 1946, Gubler et al. 1984, Beaty et al. 1989). Isolation of viral proteins - The viral proteins were extracted using polyacrylamide gel electrophoresis (PAGE) in the presence of sodium dodecyl sulfate (SDS-PAGE). Slab gels were performed with 10% separating gel and 5% stacking gel, and in both the viral proteins were revealed using 0.2% aqueous solution of commassie blue enlarged of acetic acid and ethanol (Laemmli 1970). Molecular weights of MDCV proteins were compared with five molecular weight protein markers in the same gel.
Electron microscopy Negative staining - The supernatant of Vero cells infected with MDCV was applied to grids that were covered with Formvar and a carbon film. A drop of 2% phosphotungstic acid (PTA) solution, pH 6.8, was then added. The excess of PTA was removed 30 sec later with a filter paper (Almeida 1980), and the grids were then air-dried and examined in a transmission electron microscope (TEM), model Zeiss EM 900. Ultrathin sections - Vero cells, brain and liver fragments of infected suckling mice were used for conventional electron microscopy. The specimens were fixed at room temperature for 2 h in 4% paraformaldehyde-2.5% glutaraldehyde in 0.1M cacodylate (Karnovsky 1965), pH 7.2, post-fixed with 1% OsO4 at 4°C (Hepler 1980), en bloc stained with 2.5% uranyl acetate and dehydrated through a graded series of acetone concentrations. The material was then immediately embedded in "Epon" (Luft 1961). Finally, sections were stained with uranyl acetate and lead citrate (Reynolds 1963), and observed in TEM (Zeiss EM 900). RESULTS
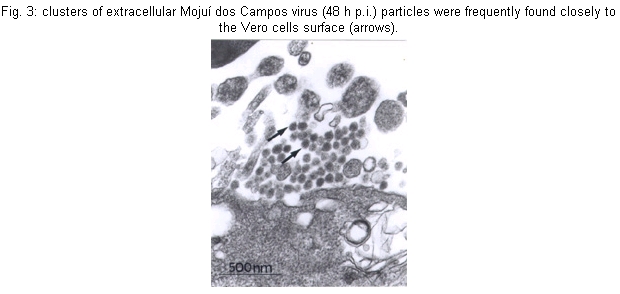
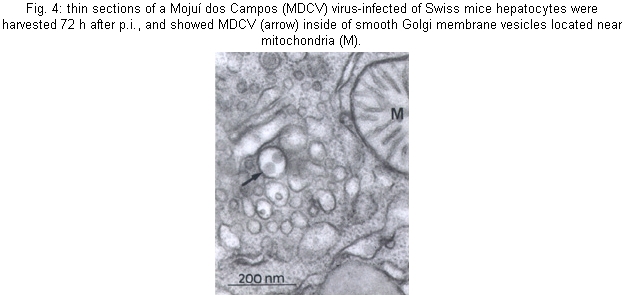
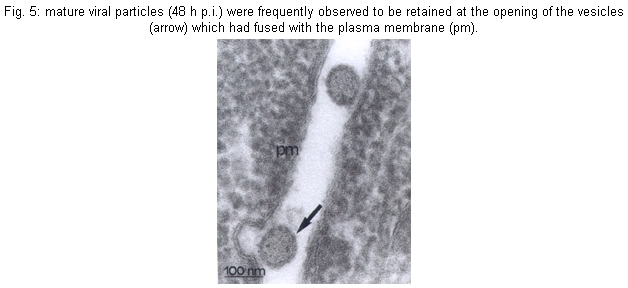
Cultures - MDCV caused CPE in Vero cells 24 h p.i., with a increasing degree of cellular destruction (Fig. 1), and at 48 h p.i. all cells were destroyed. During cultures, the morphology of the cells changed from elongated to rounded following the viral replication. Mice - Infected mice developed signs of flaccid paralysis after approximately three days, and at five days p.i. all animals had died. Physicochemical and serologic tests - MDCV showed sensitivity to DCA challenge. The control titer was 10-5.3 LD50/0.02 ml, and after treatment with DCA it was reduced to < 2.5 LD50/0.02 ml. Subsequently, MDCV was tested by the neutralization test and the logarithmic neutralization index was 10-2.6 when challenged with its homologous antiserum. In IFA tests using FICT-conjugated anti-rabbit IgG, specific fluorescence was observed in small patches in the cytoplasm (24 h p.i.) in an area immediately adjacent to and surrounding the nucleus of infected cells. Thirty hours p.i., when approximately 80% of the infected cells showed CPE, the fluorescence became more evident and generalized within the cytoplasm. MDCV showed strong reaction to the CF test only with its specific antiserum (Vero cells: ³16/³2 and brain mice: ³32/³8). Viral proteins - In order to follow the kinetics of protein synthesis, confluent Vero cell monolayers infected with MDCV and the supernatant at 3, 6, 9 and 12 h p.i. were immunoprecipitated with protein A. The MDCV proteins L, G1 and G2, displayed molecular weights of 259, 116 and 41 kDa, respectively (Fig. 1). The profile also showed presence of other proteins with molecular weights of 82 and 97 kDa. It is noteworthy that N protein was not detected in several gels made during this study. Electron microscopy - Negative stained preparations of MDCV revealed round viral particles with an average diameter of 98 nm (range: 82-100 nm). These particles had an envelope which consisted of a bilaminar membrane covered, on the surface, by projecting structures (Fig. 2). Most viral particles showed a rather irregular shape and a deformed presentation. Frequently, many of them were separated, and occasionally they occurred in groups. Ultrathin sections of infected mice brain (48 h p.i.) showed MDCV particles, especially inside the cisternae and vesicles of the Golgi structure. MDCV particles (Fig. 3) were seen forming clusters on the surface of infected cells. In the liver of infected mice at 72 h p.i., MDCV particles were observed inside the vesicles of the smooth membrane vesicles (Fig. 4). Morphological aspects of the MDCV particles in Vero cell cultures were similar to those observed in infections with other bunyaviruses (von Bonsdorff et al. 1970, Smith & Pifat 1982). Extracellular particles were seen to be round to slightly oval, with an average diameter of 83 nm (range: 70-90 nm). Virus particles were frequently found at the point of overlap between two cells (Fig. 5). DISCUSSION
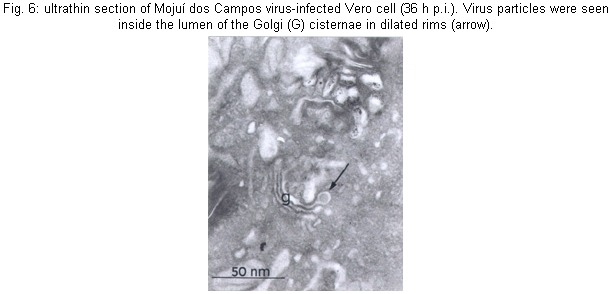
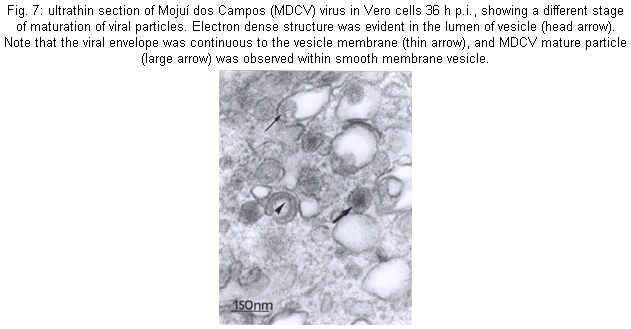
Many morphological features of the MDCV in Vero cells were similar to those previously described for other members of the Bunyaviridae (von Bonsdorff et al. 1970, von Bonsdorff & Petterson 1975, Obijeski & Murphy 1977, Patterson & Schmaljohn 1990, Gonzalez-Scarano & Nathanson 1996, Rwambo et al. 1996). The budding of the viruses Sin Nombre (Hantavirus) (Goldsmith et al. 1995) and Uukuniemi (Phlebovirus) (Kuismanen et al. 1985), occurs beneath the cell's plasma membrane, whereas the budding of MDCV, in all stages of maturation, was observed to be primarily inside membranes of the Golgi structure. Maturation in the proximity of the smooth internal membranes of the Golgi cisternae (Fig. 6) or endoplasmic reticulum has been considered to be a hallmark of members of the family Bunyaviridae (Bishop & Shope 1979, Bishop et al. 1980, Schmaljohn 1996). Development of MDCV particles was seen in all stages of replication, ranging from slightly thickened membrane crescents to essentially complete and maturated virions attached only by a thin stalk to the vesicular membrane. In sections showing budding figures, the bilaminar nature of the envelope was evident. Moreover, an electron-dense layer corresponding to condensed RPN was apparent on the cytoplasmic surface immediately under the developing envelope. Viral particles were frequently found in the lumen of the Golgi cisternae in the dilated rims (Fig. 7). It was clear that the vesicular membrane and the emerging viral envelope constitute a single continuous membrane. Similarly, an increased density was apparent on the lumenal face of developing virions, corresponding to virion spike structure. Several images indicated that the release of the virus particles from infected cells to be mediated by exocytosis. The vesicles containing mature virions were also seen close to the plasma membrane. The absence was noted of the coat on the cytoplasmic surface which is often associated with pycnocytic vesicles (Silverstein et al. 1977, Lodish et al. 1995). The study of protein synthesis eliminated any doubt regarding the classification of the MDCV as a member of Bunyaviridae. The kinetics of synthesis of MDCV-specified proteins is completely in accordance with those of other bunyaviruses of the genus Bunyavirus (Eliott et al. 1991). These findings substantiate the previous characterization proposed by Zeller et al. (1989) establishing MDCV as a member the Bunyaviridae, genus Bunyavirus. However, it remains ungrouped, since it does not share similarities with other bunyaviruses. Evidences that MDCV is by CF tests antigenically different from the other 102 arboviruses and antigenic groups isolated in the Amazon region or another region were previously described (Pinheiro & Travassos da Rosa, 1985). Moreover MDCV was also negative to Água Preta and Parixá viruses (Herpesviridae) and Mapuera virus (Paramyxoviridae), all of them also isolated from bats in the Amazon region (Pinheiro & Travassos da Rosa 1985). Brain mice infected with MDCV by CF has shown reaction only against its specific antiserum (³128/³16) and has also shown negative results to prototype of California group - California encephalitis virus, Jerry Slough, as well as to Tahyna, Snowshoe hare, Melao, Serra do Navio, Trivittatus, La Crosse, San Angelo, Keystone, Lumbo, South River, Guaroa and Inkoo. Similarly, cross reactions were not observed to the prototype of Nyando serogroup (Travassos da Rosa - pers. commun.).
ACKNOWLEDMENTS
To Eliana Vieira Pinto for help in the viral proteins techniques, and Prof. Ralph Lainson for English revision of the manuscript. REFERENCES
The following images related to this document are available:Photo images[oc02059f6.jpg] [oc02059f4.jpg] [oc02059f2.jpg] [oc02059f7.jpg] [oc02059f1.jpg] [oc02059f3.jpg] [oc02059f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}