 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(3) 2002, pp. 377-380 Effect of Antimalarial Drugs on Plasmodia Cell-Free Protein Synthesis Ana Ferreras, Ledia Triana, Erlinda Sánchez, Flor Herrera+
Centro de Investigaciones Biomédicas,
Facultad de Ciencias de la Salud, Universidad de Carabobo-Núcleo Aragua,
Apartado 2351, Maracay, Edo. Aragua, Venezuela This work was supported in part by Fundacite-Aragua, by the Consejo de Desarrollo Científico y Humanístico of the Universidad de Carabobo and by the Consejo Nacional de Investigaciones Científicas y Tecnológicas. Received 9 March 2001 Code Number: oc02072 A cell-free system from Plasmodium falciparum able to translate endogenous mRNA was used to determine the effect of artemisinin, chloroquine and primaquine on the protein synthesis mechanism of the parasite. The antimalarial drugs did not inhibit the incorporation of [3H] methionine into parasite proteins even at concentrations higher than the ones found to strongly inhibit the parasite growth. Results clearly indicate that these compounds do not have a direct effect on protein synthesis activity of P. falciparum coded by endogenous mRNA.
Key words: Plasmodium falciparum - protein synthesis - cell free translation - antimalarial drugs Several drugs are widely used alone or in combination for the treatment of falciparum malaria (Fivelman et al. 1999, Brockman et al. 2000, Nosten et al. 2000). The site of action of the majority of them is still not understood although this is a crucial information for the study of the mechanism of resistance of Plasmodium falciparum to these compounds (Warhurst 1999). There are several targets for antimalarials: the digestive vacuole, enzymes involved in membrane processes and signalling and those responsible for macromolecular and metabolite synthesis. Some of the processes taking place in the digestive vacuole are: haemoglobin digestion, redox reactions, free radical synthesis, and haem release followed by its polymerization into haemozoin. It has been reported that chloroquine (Fitch 1998, Bray et al. 1998) and artemisinin act by interfering with the heme metabolism of the parasite (Pandey et al. 1999, Padmanaban & Rangarajan 2000). The digestive vacuole may also be involved in the mechanism of chloroquine resistance (Saliba et al. 1998). Other roles have been suggested for artemisinin: alteration of membrane protein export from the vacuolar-network of P. falciparum (Akompong et al. 1999), alkylation of one or more essential malaria proteins (Meshnick 1998) and inhibition of the protein synthetic machinery of the parasite (Gu et al. 1983, Ellis et al. 1985). This last effect has also been reported for other antimalarials. Chloroquine, quinine, artemisinin, and sodium artelinate had an inhibitory effect on the [3H] hypoxantine and [3H] isoleucine uptake of cultured P. falciparum (ter Kuile et al. 1993). The effect of chloroquine may be mediated through the inhibition of the heme-dependent protein synthesis in P. falciparum (Surolia & Padmanaban 1991) concomitantly with the phosphorylation of the parasite initiation factor 2 (eIF-2). Recently, we developed a P. falciparum cell-free system that contains polyribosomes, tRNA, aminoacyl-tRNA synthetases and factors that allow the system to complete the steps of elongation and termination of the protein synthesis mechanism (Ferreras et al. 2000). Using this system we could determine that the translation of endogenous mRNA is not the target for artemisinin, primaquine and chloroquine which were highly effective inhibiting cultures of P. falciparum. MATERIALS AND METHODS
Maintenance of the malarial parasite - P. falciparum FCB1 strain, a local isolate from Colombia, was cultured in vitro according to the candle jar method (Trager & Jensen 1976). Isolation of malaria parasites from infected blood - Parasites were synchronized using 5% D-sorbitol (Freeman & Holder 1983) and the cultures were pooled at late trophozoite stage with 6-10% parasitaemia. The erythrocytes were isolated and the parasites released from them as described by Ferreras et al. (2000). Preparation of the parasite cell-free system - This was performed as described by Ferreras et al. (2000). Preparation of drug solutions - Artemisinin stock solution (8 mM) was prepared in a mixture of water and N,N-dimethyl formamide (Gu et al. 1983). Chloroquine and primaquine were dissolved in water. Then the solutions were aliquoted and stored frozen (-20°C) for no longer than one month. To establish the effect of all the drugs on parasite growth in vitro, dilutions of these compounds (0.02-200 µM) were made in complete culture medium. Assay of drug action on cultures of P. falciparum - Micro in vitro tests were performed to assess the susceptibility of P. falciparum to artemisinin, chloroquine and primaquine following the method described in Webster et al. (1985) with some minor modifications. A 190 µl aliquots of the infected erythrocytes (0.8-1% parasitemia) suspended (2% cell suspension) in growth medium (RPMI supplemented with 5% human serum of the AB type, 25 mM HEPES and g/l NaHCO3) were pipetted into each well of micro-culture plates that contained artemisinin, chloroquine, or primaquine. For each drug and the control six replicates were made. The micro-culture plates were gently agitated, placed in an airtight candle jar and incubated for 24 h at 37°C. After incubation, the contents of three replicates of each experimental tube and the control were harvested by removal of the supernatant and the red blood cells deposited on the flat bottom of the wells were transferred to a microscope slide to form thick films, which were stained for 30 min in Giemsa stain. The number of parasites was counted twice. Following the 24 h incubation, the growth medium was changed with (experimental group) or without drug (control group) to the three other group of replicates and incubation continued for 24 h more. Protein synthesis - Cell-free protein synthesis was carried out in a standard reaction mixture (Ferreras et al. 2000) containing the following components in a volume of 50 µl: 20 mM Hepes/KOH pH 7.4; 100 mM KOAc; 5 mM Mg(OAc)2; 2 mM DTT; 10% glycerol; 0.5 mM ATP; 0.1 mM GTP; 20 mM creatine phosphate; 10 µg of creatine phosphokinase; 40 µM (each) of 19 aminoacids (except methionine) and 1.7 µM L-[3H]methionine (77x 103 cpm/pmol), 140 µg of protein of S-30 fraction and variable amounts of artemisinin, chloroquine or primaquine as indicated in the legend to the Fig. 2. Reactions were initiated with the addition of the S-30 fraction and incubated at 37°C for 60 min. At the end of the incubation period the fractions insoluble in hot (90°C) 10% trichloroacetic acid were prepared; the precipitates were collected on glass-fiber filters, washed, dried and counted in a scintillation counter. Polyacrylamide gel electrophoresis - The proteins were analyzed by 12.5% SDS/PAGE according to Laemmli (1970). Some incubations for protein synthesis were performed using [35S] methionine (117,5 Ci/mmol). Radioactively labeled proteins were electrophoresed and visualized by autoradiography. Protein concentration determination - Protein concentration was determined as described by Lowry et al. (1951). RESULTS
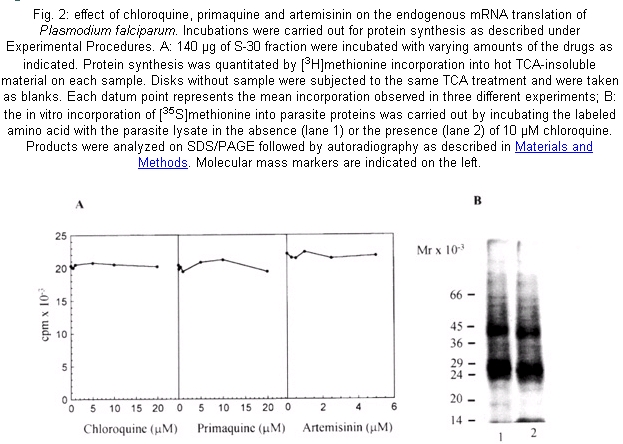
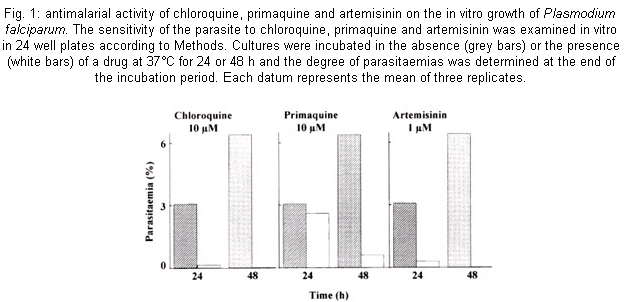
The susceptibility of cultures of P. falciparum to chloroquine, primaquine and artemisinin was studied by subjecting them to different concentrations of the drugs (0.02 -200 µM) throughout the 48-h erythrocytic life cycle. Fig. 1 shows that exposure for 24 h of infected erythrocytes to chloroquine or primaquine at 10 µM and artemisinin to a concentration of 1 µM caused an inhibition of parasite growth of 95%, 15% and 90% respectively. The inhibition due to primaquine increased to 90% following the incubation up to 48 h; while artemisinin and chloroquine produced a 100% growth inhibition. The effects of these compounds were examined in a cell-free system from P. falciparum that we developed very recently (Ferreras et al. 2000). This system is able to translate endogenous mRNA giving about 25-fold stimulation over the activity at time zero which indicates that the parasite cell-free system contains polyribosomes, tRNA, aminoacyltRNA synthetases and factors that allow the system to complete the steps of elongation and termination of the protein synthesis mechanism. The drugs did not inhibit the incorporation of [3H] methionine into parasite proteins even at concentrations higher than those found to strongly inhibit the parasite growth (Fig. 2A). Also, the autoradiograph of the proteins synthesized by the parasite cell-free system clearly shows polypeptides ranging from 66 kDa to 24 kDa (Fig. 2B, lane 1). A similar result was observed even in the presence of 10 µM chloroquine in the system (Fig. 2B, lane 2). Comparable data were obtained with primaquine and artemisinin (data not shown). DISCUSSION
The main objective of this work was to ascertain if artemisinin, chloroquine and primaquine could cause an inhibitory effect on the endogenous mRNA translation of the P. falciparum since it has been reported that some antimalarial drugs inhibit parasite protein synthesis (Gu et al. 1983, Surolia & Padmanaban 1991, ter Kuile et al. 1993). Our strategy was firstly to find the concentrations of the drugs that inhibit parasite growth drastically. Secondly, it was determined if these lethal drug concentrations could act similarly on the elongation of P. falciparum protein synthesis. We found that chloroquine and primaquine at 10 µM and artemisinin at 1 µM (Fig. 1) caused a sharp decline in parasitemia, a drastic inhibition of parasite growth within a single intraerythrocytic asexual cycle (approximately 48 h). However, the drugs were not effective against the mechanism of endogenous protein synthesis in the plasmodia cell-free system irrespective of the high concentrations used. Indeed, they did not change the pattern of the proteins synthesized (Fig. 2). These results clearly reveal that chloroquine, primaquine and artemisinin do not have a direct effect on the elongation step of the protein synthesis activity of P. falciparum. Other reports suggested that the drugs exert their effects by targetting the protein synthesis mechanism of P. falciparum (Gu et al. 1983, Surolia & Padmanaban 1991, ter Kuile et al. 1993). These findings do not contradict our results since we tested the effect of drugs on the endogenous mRNA translation that mainly represents the elongation step of the protein synthesis mechanism. It still remains to be elucitated whether the drugs could inhibit the initiation event of the process. Another possibility is that the earliest action of the drugs may cause other defects ultimately leading to a decrease in general polypeptide synthesis of the parasite. Thus, this cell-free system can be used as a tool to evaluate the effect of antimalarial agents in the endogenous mRNA translation of P. falciparum.
ACKNOWLEDGEMENT
To Drs Edmundo Cayama and Ricardo Montoreano for helpful discussions and to Maria Briceño for culturing parasites. REFERENCES
The following images related to this document are available:Photo images[oc02072f1.jpg] [oc02072f2.jpg] |
| |||||||||

{kind=link}
{kind=link}