 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(3) 2002, pp. 411-414 Prevalence of Trypanosoma lewisi in Rattus norvegicus from Belo Horizonte, State of Minas Gerais, Brazil Pedro Marcos Linardi+, José Ramiro Botelho
Departamento de Parasitologia, Instituto
de Ciências Biológicas, Universidade Federal de Minas Gerais, Caixa
Postal 486, Received 27 March 2001 Code Number: oc02078
From April 1984 to March 1985, a Trypanosoma lewisi prevalence of 21.7% was found in 429 Rattus norvegicus trapped in Belo Horizonte, State of Minas Gerais, Brazil. The infection rates were higher in male and young rats and could be attributed to ecological and behavioral factors. T. lewisi was observed in rats measuring between 60 and 250 mm. Data about monthly T. lewisi infections throughout the year are presented for the first time in Brazil, with the highest prevalences observed in the warm-rainy season (October to March).
Key words: Trypanosoma lewisi - blood trypomastigotes - Rattus norvegicus - infection rates - sex-age host - seasonal periods - Minas Gerais - Brazil The trypanosomes of mammals are divided into two sections, Stercoraria and Salivaria, according to the developmental cycle in vectors (Hoare 1972). In Stercoraria, the development is completed in the faecal medium of the posterior portion of the insect and transmission is contaminative. One of the subgenera included in this section, Herpetosoma, comprises about 45 species, whose adult blood forms are morphologically indistinguishable from those of the type-species, Trypanosoma (Herpetosoma) lewisi. This species parasitizes synanthropic rodents of the genus Rattus and has rat-fleas as vectors. Although Xenopsylla cheopis, Nosopsyllus fasciatus, Ctenocephalides canis and C. felis have been incriminated as intermediate hosts (Molyneux 1969), X. cheopis is the principal vector in tropical and subtropical areas (Hoare 1972). Although this trypanosome is a non-pathogenic blood flagellate, Guerrero et al. (1997) demonstrated that T. lewisi infection increases Toxoplasma gondii multiplication in white rats. Studies on prevalence of T. lewisi in murine populations were mainly made at the middle of the century by Laird (1951) in New Zealand, Callero (1952) in Panama, Eyles (1952) in USA and Kartman (1954) in Hawaii. More recently, this infection has been reported from Asiatic or African countries by Linxian et al. (1994) in the People's Republic of China, Ugbomoiko (1997) in Nigeria, Laha et al. (1997) in India, and Abdel-Aal and Abou-Eisha (1997) in Egypt. Up to now, South American records include Chile (Franjola et al. 1995) and Venezuela (Herrera & Urdaneta-Morales 1997). Here we report the prevalence of T. lewisi in connection with sex and host age in Rattus norvegicus and the monthly examination records for Brazil. MATERIALS AND METHODS
During a rat-flea survey for epidemiological surveillance in Belo Horizonte, State of Minas Gerais, Brazil, 429 R. norvegicus were trapped around dumps at the margins and surroundings of the Rio Arrudas, from April 1984 to March 1985. Rats were killed by chloroforming and blood from the tail was used to prepare the blood films later stained with Giemsa. Trypanosomes were identified after examination under a 100x objective. Rats were separated according to sex and divided into six age-groups considering the length of the head plus body from 0 to 280 mm, and were also classified as immatures (0 to 140 mm); young matures (141 to 200 mm) and adults (more than 201 mm). Monthly prevalences were also determined and grouped by seasonal periods, with a dry-cool season from April to September 1984 and a warm-rainy season from October 1984 to March 1985. The chi-square analysis was used to determine the degree of association between infection and factors considered. RESULTS
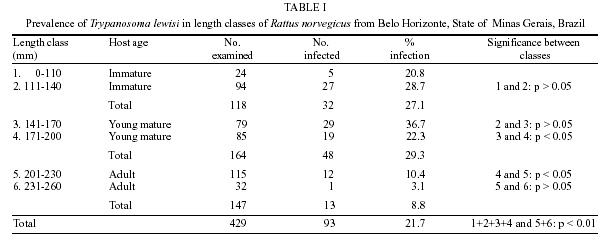
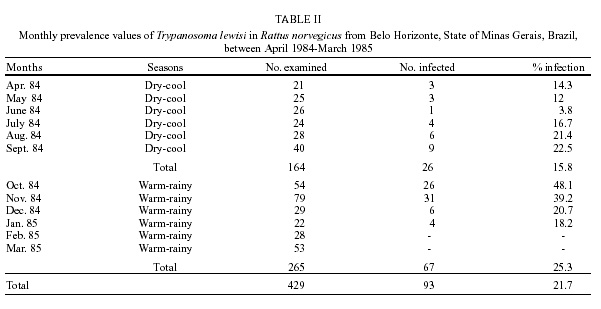
T. lewisi was observed in a total of 93 out of 429 R. norvegicus (21.7%). The infection rate was different in both sexes, 24.9% (68/273) being detected for males and 16% (25/156) for females. The infection rates related to length and host age are shown in Table I. The smallest rat in which T. lewisi was found measured 60 mm and the largest one measured 250 mm. The monthly prevalence and the infection rates of T. lewisi related to seasonal periods are shown in Table II. DISCUSSION
The overall prevalence of rats found to be infected with T. lewisi in the present study (21.7%) is higher than that for any other country except India, where a value of 82.3% was recorded (Laha et al. 1997). Other studies have recorded values of 4.6% in New Zealand (Laird 1951), 8.9% in Nigeria (Ugbomoiko 1997), 11.4% in Hawaii (Kartman 1954), 13.2% in both the USA (Eyles 1952) and Egypt (Abdel-Aal & Abou-Eisha 1997), 20% in Italy (De Carnieri & Castellino 1964) and 21.3% in Venezuela (Herrera & Urdaneta-Morales 1997). Unlike in the studies of Franjola et al. (1995) and Ugbomoiko (1997) there was a significant difference (p < 0.05) between the infection rates in males (24.9%) and females (16%). These higher prevalences could be attributed to ecological and behavioral factors. Male rats have larger home ranges, show territorial behavior and are significantly more infested by X. cheopis than females (Linardi et al. 1985a), increasing their chances of being infected by T. lewisi infections. Different age-related rates of infection have been recorded worldwide. Although Franjola et al. (1995) found no significant differences among age groups and Ugbomoiko (1997) observed higher values in adult rats, in our study the prevalence of infection in young animals (29.3%) was similar to that of immatures (27.1%) and more than three times higher that that recorded for adults (8.8%). These differences are highly significant (Table I) and agree with the results of Eyles (1952) and Kartman (1954). The infection rates gradually increased with total body length in animals measuring 60-170 mm and declined thereafter. The highest rates were found in young animals (141-170 mm) and immatures (111-140 mm). Significant differences were found between the infection rates of length classes 3 (141-170 mm) and 4 (171-200 mm) as well as between classes 4 and 5 (201-230 mm). The profile of infection in T. lewisi is well known (Albright & Albright 1991). After a rapid multiplication for 10 days, the trypanosomes stop growing and infection stabilizes for several weeks, after which the parasites suddenly disappear from the blood and the rat develops a solid immunity against re-infection. The trick used by T. lewisi to minimize the effects of the immune responses of the host is known as mimicry. Initially the parasites coat their surfaces with ablastin, a host protein later identified as immunoglobulin E, that prevents the trypanosomes from dividing and multiplying. After several weeks, the host produces an IgM antibody that recognizes this complex on the parasite's surface and leads to the activation of the complement system resulting in rapid lysis of the trypanosomes in the blood. For this reason blood smears from adult rats show fewer trypanosomes than those from young or immature animals. Since norway rats attain sexual maturity at 75 days and only 5% of the population survives 12 months (Davis 1948), the animals probably become infected before or during their first mating. Use of hemoculture rather than examination of blood films would show infections in many rats, mainly adults, that would otherwise be considered negative. Trypanosomes were found throughout the year except in February and March (Table II). Rats presented significantly higher prevalences of infection (p < 0.05) in the warm, rainy season (October to March) than in the cool, dry period (April to September). The highest values, recorded in October (48.1%) and November (39.2%) coincide with highest levels of infestation by X. cheopis in the municipality (Linardi et al. 1985a,b). Infection rates increased from June to October and subsequently decreased. Infection rates of T. lewisi in Brazilian commensal rats are presented for the first time. Although this study provided data from only a single year, they show that infection rates are influenced by climatic conditions that affect the size of flea populations (Bahmanyar & Cavanaugh 1976, Gratz & Brown 1983, Linardi et al. 1985b). Guerrero et al. (1997) and Arrea et al. (1998) reported that under experimental conditions T. lewisi increased T. gondii multiplication in white rats, changing the susceptibility of the host to the parasite. This observation deserves greater attention than it received when published, since (a) T. gondii-infected rats are considered important in the epidemiology of toxoplasmosis because they can serve as reservoirs of infection for pigs, dogs and cats (Dubey & Frenkel 1998); (b) a high prevalence of T. gondii was found recently in R. norvegicus on farms in England, suggesting that Toxoplasma infections can be perpetuated in wild rodent populations without the presence of cats (Webster 1994); (c) T. gondii infection may enhance the likelihood of infected rats being predated by cats (Webster 1994, Berdoy et al. 1995a,b). Since both T. lewisi and T. gondii occur naturally in wild rats, it is possible that infections by the former could increase the number of cysts in the brain, making these rodents important sources of Toxoplasma infection for cats (Arrea et al. 1998). Thus, if increasing levels of infestation of rats by X. cheopis favor T. lewisi transmission, which in turn promotes the spread of T. gondii, control measures against rat toxoplasmosis should be considered. The degree to which peaks in flea population are linked to higher prevalences of T. gondii in rats needs to be elucidated, to determine whether flea control might result in control of rat toxoplasmosis. REFERENCES
© 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02078t2.jpg] [oc02078t1.jpg] |
| |||||||||

{kind=link}
{kind=link}