 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(4) 2002, pp. 553-557 In Vitro and in Vivo Anti-Trypanosoma cruzi Activity of a Novel Nitro-derivative Susana Muelas-Serrano, Ana Le-Senne,
Carlos Fernández-Portillo, Juan José Nogal, Carmen Ochoa*,
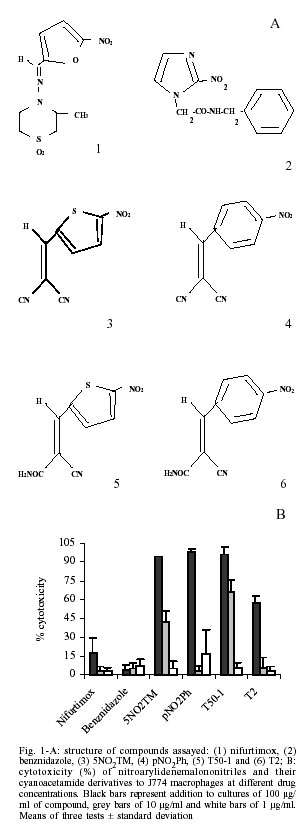
Alicia Gómez-Barrio/+ Received 10 September 2001 Code Number: oc02105 Departamento de Parasitología, Facultad de Farmacia, Universidad Complutense, Ciudad Universitaria s/n, 28040 Madrid, España *Instituto de Química Médica, Consejo Superior de Investigaciones Científicas, Madrid, España Nitroarylidenemalononitriles and their cyanoacetamide derivatives with remarkable anti-epimastigote properties, were synthesized attempting to obtain new 3,5-diamino-4-(5'-nitroarylidene)-4H-thiadiazine 1,1-dioxide derivatives, which in previous reports had shown anti-Trypanosoma cruzi activity. Tests to evaluate the cytotoxicity of compounds were performed on J774 macrophages. 5-nitro-2-thienyl-malononitrile (5NO2TM), was the only product which maintained a high anti-epimastigote activity at concentrations in which it was no longer cytotoxic, thus it was assayed against intracellular amastigotes. Its anti-amastigote activity was similar to that of nifurtimox. Afterwards in vivo toxicity and anti-chagasic activity were determined. A reduction in parasitemia was observed. Key words: Trypanosoma cruzi - nitroarylidene - malononitriles - in vitro assays - cytotoxicity assays - in vivo experiments Chagas disease is endemic throughout Latin America. It is estimated that 16-18 million people are infected by Trypanosoma cruzi (WHO 1993), the causative agent of this disease. Lately, great advances have been made in the control of vectorial and transfusion transmission of the disease, through spraying programs to eliminate the triatomine vectors and screening of blood banks (TDR 2000). Nevertheless, since the discovery of this disease more than 90 years ago, there is still no efficient treatment. A large number of different compounds have been assayed in a variety of ways, for instance cysteine-protease inhibitors that recognize cruzipain active site are effective in acute and chronic murine models (Engel et al. 1998): phenothiazines inhibit trypanothione reductase, acting in this way as promising trypanocidal agents (Chan et al. 1998). In spite of the amount of work conducted in studying trypanothione reductase and proteases in particular, still the drugs available for clinical use are restricted to benznidazole and nifurtimox (de Castro 1993). Both can reduce symptoms and mortality in the acute phase of the illness, but are not effective in achieving parasitologic cure or preventing the chronic phase. In addition, both drugs are highly toxic (Jernigan & Pearson 1993). Although there is an important number of publications concerning the development of new chemotherapeutical agents and target-based drugs against T. cruzi, such as sterol and phospholipid biosynthesis (Urbina et al. 1996, Abt et al. 1998), purine salvage pathway (Eakin et al. 1997), tubulin and trypanosome transporters inhibitors (Werbovetz 2000), the search for effective treatment for Chagas disease has been met with limited success. It remains, thus, an open field of investigation. Previous works reported the antichagasic properties of 3,5-diamino-4-(5'-nitroarylidene)-4H-thiadiazine-1,1-di oxides (Atienza et al. 1992, Herrero et al. 1992). Attempts to synthesize new compounds with this structure failed and only the cyanoacetamide derivatives were obtained by partial hydrolysis of one of the nitrile groups from 5-nitro-arylidenemalononitriles (di Maio et al. 1999). The anti-epimastigote activity of these compounds was then evaluated and it was noteworthy that the activity of 5-nitro-thienyl-malononitrile (5NO2TM) was higher than nifurtimox activity (Muelas-Serrano et al. 2001a, b). The interesting activity of these new nitro-derivatives without the tetrahydrothiazine moiety of nifurtimox encouraged us to carry out new experiments to study in depth the properties of these compounds. MATERIALS AND METHODS Cell culture - Murine J774 macrophages were grown in plastic 25 µl flasks in RPMI 1640 medium (Sigma) suplemented with 20% heat inactivated (30 min, 56oC) foetal calf serum (FCS) and 100 IU penicillin/ml + 100 µg/ml streptomycin, in a humidified 5% CO2/95% air atmosphere at 37oC and subpassaged once a week. Parasites - Trypanosoma cruzi Chagas, 1909 (Y strain) was grown at 28oC in liver infusion tryptose (LIT) supplemented with 10% FCS and antibiotics. Epimastigote forms were harvested on day 14 of culture (stationary phase) and washed three times in Grace medium. To induce metacyclogenesis, parasites were then cultured in fresh Grace medium supplemented with 10% FCS and haemin (25 µg/ml). Nine days after cultivation at 28oC, metacyclic forms were counted. The proportion of metacyclic forms was around 30% at this stage. These metacyclic forms were used to infect J774 macrophages, for anti-amastigote assays. Cell infection - J774 macrophages were detached by EDTA-PBS (ethylendiamine tetraacetic acid- phosphate-buffered saline) treatment and counted in a hae-mocytometer. Cells were seeded at a density of 50,000 cells/well in 24-well microplates (NUNC) with rounded coverslips on the bottom. Then 500,000 metacyclic trypomastigotes and fresh medium were added, giving a final volume of 2 ml. Attachment and invasion of host cells were allowed for 24 h. Cytotoxicity to macrophages - J774 macrophages were seeded (70,000 cells/well) in 96-well flat bottom microplates (NUNC) with 200 µl of medium. The cells were allowed to attach for 24 h at 37ºC and then exposed to the compounds (100, 10 and 1 µg/ml, dissolved in DMSO, maximum final concentration of solvent was 0.2%) for another 24 h. Afterwards, the cells were washed with PBS and incubated (37ºC) with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) 0.4 mg/ml for 60 min. MTT solution was removed and the cells solubilized in dimethyl sulphoxide (100 µl). The extent of reduction of MTT to formazan within cells was quantified by measurement of OD595 (Hattori & Nakanishi 1995). Each concentration was assayed three times and six cell growth controls were used in each test. The assays were performed twice. Nifurtimox and benznidazole cytotoxicity was also determined. Cytotoxicity percentages (%C) were determined as follows:
%C = [1-(ODp-ODpm)/(ODc-ODm)]x100
where ODp represents the mean OD595 value recorded for wells with macrophages containing different doses of product; ODpm represents the mean OD595 value recorded for different concentrations of product in medium; ODc represents the mean OD595 value recorded for wells with macrophages and no product (growth controls), and ODm represents the mean OD595 value recorded for medium/control wells. The cytotoxic dose 50 (CD50) was defined as the concentration of drug which decreases OD595 up to 50% of that in control cultures. Anti-amastigote activity - After cell infection, culture medium was removed to eliminate non-infecting trypanosomes, and suspensions of compound 5NO2TM in fresh medium were added to final concentrations non-toxic for macrophages (10, 1 and 0.5 µg/ml). 5NO2TM was the only product, out of the four, that remained active against epimastigotes at concentrations non-toxic for macrophages. After 48 h, the coverslips were fixed and stained with May Grünwald Giemsa and the number of amastigotes/100 macrophages (No. A/100 Mø) were estimated. Anti-amastigote activity (%AA) was expressed as:
%AA = [1- (No. A/100 Mø)p/(No. A/100 Mø)c]x100
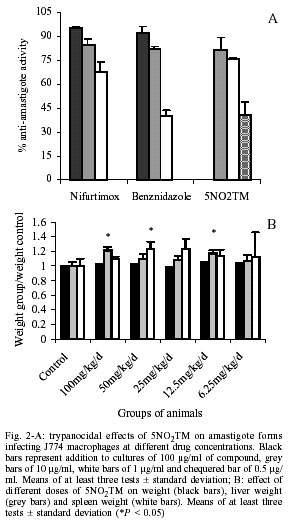
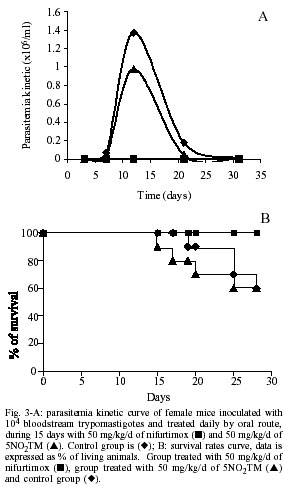
All experiments were run at least in triplicate and the results are given as mean ± standard deviation (Mendez et al. 1999). Nifurtimox and benznidazole were used as reference drugs. In vivo toxicity - For in vivo experiments, NMRI female mice of 45 days (at the beginning of test), weighing 20-22 g, were used. Product was dissolved in 2% carboxymethyl cellulose, doses that ranged from 6.25 to 100 mg/kg were given daily by gavage for 15 days. For each treatment a group of 6 animals was used. When treatment finished, all the animals were weighed and four of each group sacrificed to examine lesions on liver and spleen, alterations in weight and colour were recorded. The other two animals were left to verify that no death took place after treatment. Statistical analysis was performed using the non-parametric Mann-Whitney U test to compare lesions of experimental and control groups of mice. Anti-Trypanosoma in vivo test - Infected blood was obtained from anaesthetised donor-animals by intra-cardiac puncture, and diluted with non-infected blood yielding the desired number of blood trypomastigotes. NMRI female mice were infected by intraperitoneal injection of 104 blood trypomastigotes. Three groups of 10 animals were prepared. One was an infection control, the second was treated with nifurtimox 50 mg/kg/d given by gavage for 15 days and the third was administrated the new compound. The treatment was initiated 3 days after infection and maintained during 15 days. The level of parasitemia was checked by counting in a Neubauer Chamber the number of parasites in 5 µl of blood drawn from the tail of the mice and diluted 1:10 in ammonium chloride (Barr et al. 1995). The mortality was noted daily and the percent of survival calculated. Source of compounds - The synthesis of the 4 new compounds, nitroarylidenemalononitriles and their cyanoacetamide derivatives, was described elsewhere (di Maio et al. 1999). Nifurtimox (Lampit; Bayer, Buenos Aires, Argentina) and benznidazole (Rochagan; Roche, Rio de Janeiro, Brazil) were used as the reference drug in every assay (Fig. 1A). RESULTS Cytotoxicity properties of nitroarylidenemalononitriles and their cyanoacetamide derivatives were evaluated against J774 macrophages at the doses of 100, 10 and 1 µg/ml. The toxicity percentages found are presented in Fig. 1B. All the new compounds showed high toxicity at 100 µg/ml, while at 10 µg/ml they were no longer toxic, except for product T50-1. Nevertheless, at this and lower concentrations only 5NO2TM remained active against epimastigotes, thus it was the only product assayed against amastigotes. Fig. 2A shows the activity of the new drug against amastigotes infecting J774 macrophages, compared to that of reference drugs. 5NO2TM has the same effect on amastigotes infecting macrophages than nifurtimox as 10 and 1 µg/ml. At 0.5 µg/ml it maintained the same activity as benznidazole at twice this concentration. The first in vivo assay was performed to determine the toxicity of this product for mice. 5NO2TM showed a relatively high toxicity against Vero fibroblasts (di Maio et al. 1999), so with this assay we tried to ensure a dosage for the treatment of animals that would guarantee their safety and not cause death. This assay is not a LD50, but it assures us that when animals are treated with this same pattern of doses, no damage is derived from the administration of the compound during the period of time that acute infection lasts. Doses administrated ranged from 6.25 to 100 mg/kg/d. None of the animals died from treatment after 60 days. However hepatic lesions could be observed after treatment with 100 mg/kg/d. None of the other parameters studied were of significant relevance (Fig. 2B). Taking into account these data we preceded with the evaluation of the effect of 50 mg/kg/d of 5NO2TM on mice infected with T. cruzi. The oral administration of this compound to mice inoculated with 104 parasites caused a 30% decrease in parasitemia when compared with controls (Fig. 3A), but the recorded activity was not as remarkable as the one from nifurtimox. Mortality of 5NO2TM treated and untreated groups is the same (Fig. 3B), while animals treated with reference drug did not die. DISCUSSION T. cruzi is a protozoan parasite whose life cycle involves invertebrate and vertebrate hosts. Epimastigotes are the extracellular multiplying form of the parasite in the reduvid insect-vector. The fact that this form of the parasite can be easily cultured and that is relatively sensitive to drug action makes this model an excellent one for preliminary in vitro screening. However, once these first assays have been performed, more selective methods are required to determine the activity of new compounds. In this sense non-specific cytotoxicity assays are needed to assure that anti-protozoal activity is due to a specific mechanism against this parasite. We have developed a method to determine toxicity against J774 macrophages, cells that will be used subsequently as host for the intracellular form of the parasite. We have demonstrated that although malononitrile derivatives were active against epimastigotes (di Maio et al. 1999), this activity was mainly due to a non-specific toxicity. Products pNO2Ph, T50-1 and T2 anti-epimastigote activity decrease parallel to their cytotoxicity against macrophages. Only compound 5NO2TM remained active against T. cruzi at concentrations in which it was no longer toxic (%C < 50%), nevertheless this product was more cytotoxic than reference drugs (Fig. 1B). Multiplicative forms of the parasite that appear in humans are amastigotes. These forms proliferate intracellularly. Potential anti-chagasic drugs are to be effective against amastigotes. There are several in vitro methods for evaluation of anti-amastigote activity. While penetrating through the skin the first cells encountered by the parasite in which to multiply are macrophages, thus we have chosen these cells as the in vitro host. We treated T. cruzi-infected J774 macrophages with 5NO2TM, nifurtimox and benznidazole. A certain percentage of amastigotes are never killed by any of the drugs, but activity of the new compound is high at 10 and 1 µg/ml, demonstrating that it is as effective as reference drugs against the parasite even when it is in an intracellular location. Oral administration of this product to infected mice had also an effect on parasitemia, reducing it, but not to a level in which it caused no mortality. Survival rates were the same as for the control groups. Although we have used in this report the same doses of 5NO2TM and nifurtimox, and in vitro they achieved the same efficacy at similar concentrations, no important activity on experimentally infected mice was found. Since product was absorbed after oral administration, as demonstrated by the liver lesions caused at higher concentrations, the reason for this difference could be a rapid metabolization of 5NO2TM or a lack of suitable pharmacological properties for the in vivo treatment. The mechanism of action of 5NO2TM is not completely known, however a similar one to other nitro-derivatives should be accounted for. Nitroderivatives have been reported to have different mechanisms of action, that include: (a) nitro-anion radical direct action upon macromolecules, such as DNA (Kedderis & Miwa 1988); (b) oxiradicals generation, which are the main toxic agent in the case of nifurtimox (Docampo & Stoppani 1979); (c) production of nitrous and hydroxylamine derivatives, which possess a high cytotoxicity as well (Edwards 1993); and (d) direct inhibition of trypanothione reductase (Henderson et al. 1988), that bound to other enzymes is an anti-oxidant system against free radicals, peroxides and hydroperoxides. Nevertheless, differences observed in previous studies on the effect of this product and nifurtimox on HSP60 levels concluded that data obtained may implicate that the mode of action could differ in spite of the structure similarities of both drugs (Muelas-Serrano et al. 2001a). In vitro ultrastructural damages by 5NO2TM are greater than those caused by nifurtimox (Muelas-Serrano et al. 2001b). After the initial promising results, non-satisfactory in vivo results were found for unknown reasons, indicating that another approach must be attempted, such as intra-peritoneal administration or a longer duration of treatment. On the other hand, the facts that 5NO2TM is highly active against culture forms and in vitro effect of other thiadiazines against blood forms has been previously demonstrated (Herrero et al. 1992), induce us to believe that this compound may turn out to be an interesting chemoprophylactic drug. Chemoprophylactic tests, based on an initial study of compound activity on infected stored blood, and posterior inoculation in receptive animals, could be performed in order to assess its efficacy (Gómez-Barrio et al. 1997). REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02105f2.jpg] [oc02105f3.jpg] [oc02105f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}