 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(5) July 2002, pp. 703-710 Fibrogenesis and Collagen Resorption in the Heart and Skeletal Muscle of Calomys callosus Experimentally Infected with Trypanosoma cruzi: Immunohistochemical Identification of Extracellular Matrix Components Isis F Magalhães-Santos, Elianita S Lima, Sonia G Andrade+ Laboratório de
Doença de Chagas Experimental, Centro de Pesquisas Gonçalo Moniz-Fiocruz,
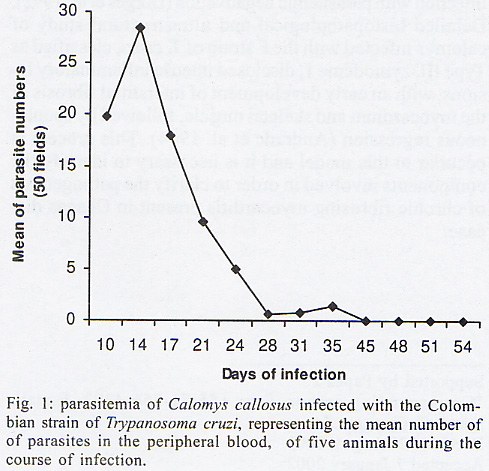
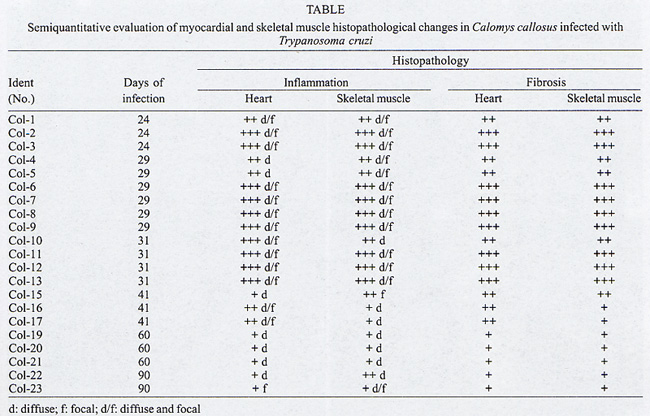
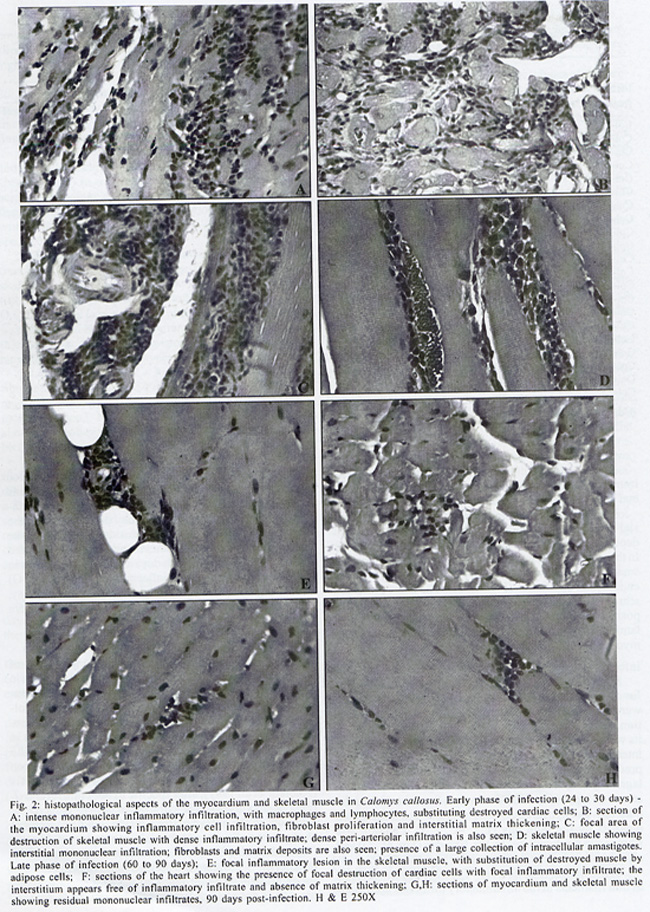
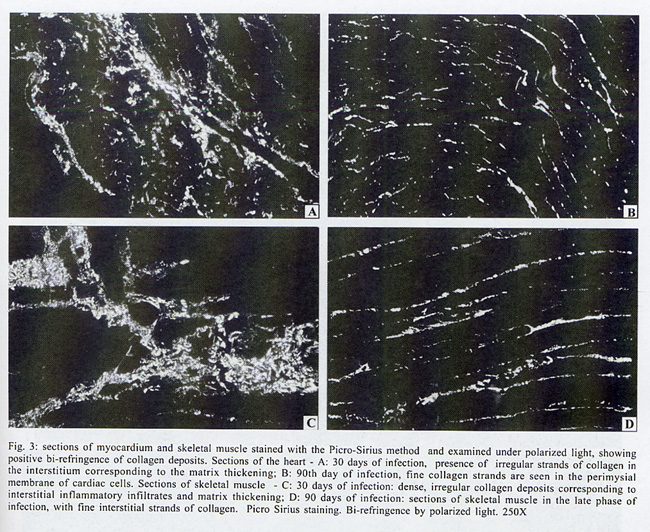
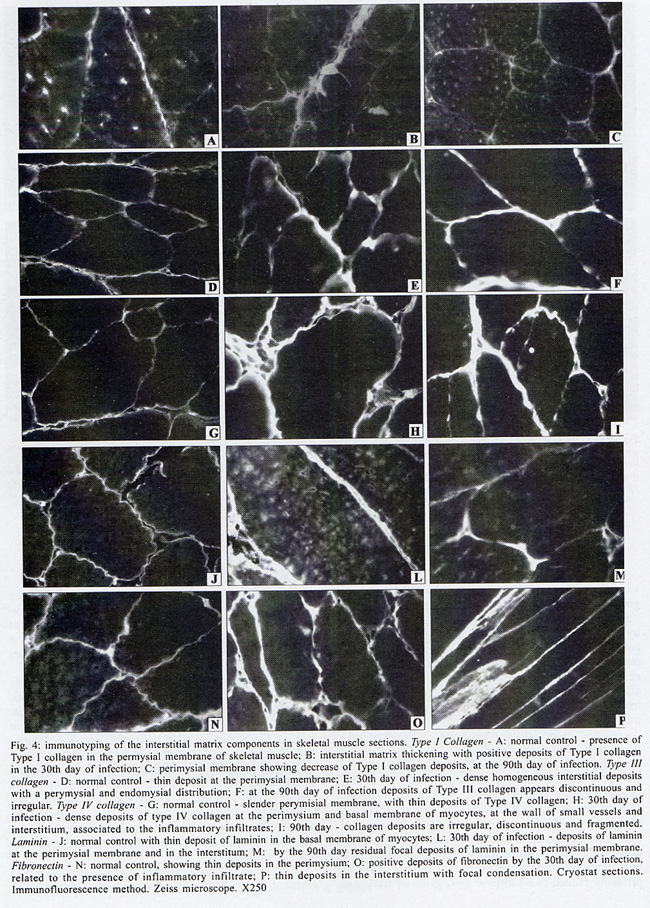
Rua Valdemar Falcão 121, Supported by Papes 2. Received 28 August 2001 Code Number: oc02135 Intense inflammatory lesions and early development of interstitial fibrosis of the myocardium and skeletal muscle with spontaneous regression, have been described in Calomys callosus infected with Trypanosoma cruzi. The genetic types of collagen present in this model were investigated through immunohistochemistry using specific antibodies, combined with histopathology and Picro-Sirius staining of collagen. Thirty-five calomys were infected with the Colombian strain of T. cruzi and sacrificed at 24, 30, 40, 60 and 90 days post-infection. Inflammatory lesions and fibrogenesis were prominent at the early phase of infection and significantly decreased during late infection. Immunoisotyping of the matrix components was performed by indirect immunofluorescence on 5 µm thick cryostat sections using specific antibodies against laminin, fibronectin and isotypes I, III and IV of collagen. In the early phase, positive deposits of all the matrix components were present, with predominance of fibronectin, laminin and collagens types I and III in the myocardium and of types III and IV in the skeletal muscles. From the 40th day, type IV collagen predominates in the heart. At the late phase of infection (60th to 90th day), a clear fragmentation and decrease of all the matrix components were detected. Findings of the present study indicate that a modulation of the inflammatory process occurs in the model of C. callosus, leading to spontaneous regression of fibrosis independent of the genetic types of collagen involved in this process. Key words: Trypanosoma cruzi - Calomys callosus - myocardial fibrosis - collagen immunotyping - interstitial matrix Calomys callosus (Rodentia cricetidae) is a wild rodent that has been found naturally infected with different pathogens such as Machupo virus (Justines & Johnson 1969) and Trypanosoma cruzi (Ribeiro 1973, Mello 1982). Several studies have been developed using C. callosus as a model for the experimental studies of Chagas disease, and its response to inoculation with different strains of T. cruzi (Borges & Mello 1980, Borges et al. 1982, 1992, Andrade et al. 1994). This animal is more resistant than mice to infection with T. cruzi, apparently controlling the infection with parasitemic negativation (Borges et al. 1992). Detailed histopathological and ultrastructural study of calomys infected with the F strain of T. cruzi, classified as Type III, zymodeme 1, disclosed intense inflammatory lesions, with an early development of interstitial fibrosis of the myocardium and skeletal muscle, followed by spontaneous regression (Andrade et al. 1994). This process is peculiar to this model and it is necessary to identify the components involved in order to clarify the pathogenesis of chronic fibrosing myocarditis present in Chagas disease. Fibrosis is one of the most important features of chronic Chagas cardiomyopathy. According to Weber et al. (1993), fibrillar collagen deposits in the myocardial interstitium are of paramount importance in the development of cardiac failure, and particularly of diastolic disfunction, in different cardiac diseases. The possibility of regression of fibrosis, already documented in the chronic infection of mice under chemotherapeutic treatment (Andrade et al. 1991) has been described as spontaneously occurring in C. callosus, probably due to down regulation of delayed type hypersensitivity (Andrade et al. 1994). However, other peculiarities of the present model must be investigated, including the sensitivity of the interstitial matrix components to spontaneous regression, regarding several types of collagen. Besides other factors such as inflammatory response and cytokines, the genetic pattern of the matrix components, which is peculiar for different species, can be responsible for the characteristic evolution of fibrosis in this model. With the objective of evaluating the participation of different genetic types of collagens in the development and regression of fibrosis in C. callosus infected with T. cruzi, an immunohistochemical characterization of the matrix components was performed, combined with an evolutive histopathological study. MATERIALS AND METHODS Experimental animals - Thirty-five C. callosus were used in this study: 30 were infected with the Colombian strain of T. cruzi and 5 were maintained as normal controls. Inoculum - 105 trypomastigotes forms were obtained from the blood of infected mice, washed three times in PBS by centrifugation and inoculated intraperitoneally. Parasitemia - Was evaluated twice per week by microscopic examination of peripheral blood under coverslips (22 x 22 mm) by counting the parasite numbers in 50 high power (x 400) fields. Histopathology - Three calomys were sacrificed at 24, 30, 40, 60 and 90 days post-infection for histopathological study. Fragments of the heart and skeletal muscle were fixed in formalin, paraffin-embedded and 5 µm-thick sections were stained with hematoxylin and eosin (H & E). Picro-Sirius staining - Paraffin sections of the heart and skeletal muscle were stained with Rot-Sirius 3 (Chroma Gessellschaft) in a saturated aqueous solution of picric acid and fast green as a counterstain to evaluate collagen deposits (Junqueira et al. 1979). Sections were examined with bright field light optics using a Zeiss photo microscope. Immunohistochemistry - For immunoisotyping of extracellular matrix components, sections of the heart and skeletal muscles from infected calomys and normal controls were removed, placed immediately in tissue Teck II O.C.T. and snap frozen in liquid nitrogen. Immunolabelling was performed by indirect immunofluorescence on 4 µm thick cryostat sections using specific antibodies against laminin, fibronectin and isotypes I, III and IV of collagens. Specific antibodies - Rabbit serum anti-mouse fibronectin, NOVOTEC-54951, lot: 01B; rabbit IgG anti-mouse laminin, NOVOTEC-24851 lot: 231; Rabbit IgG anti-mouse collagen I, NOVOTEC-220151, lot: 247; rabbit IgG anti- mouse collagen III, NOVOTEC-20351, lot: 65 and rabbit IgG anti-mouse collagen IV, NOVOTEC-20451, lot: 240. As secondary antibody, a goat anti-rabbit IgG (whole molecule) FITC conjugated (Sigma Laboratories) in the dilution of 1:10 was used. Examinations were performed in a Zeiss fluorescence microscope. For negative controls, sections of heart and skeletal muscle from infected animals and normal controls were submitted to all the steps of the reaction excluding the addition of antibodies against specific extracellular components. RESULTS In infected animals parasitemia peaked by the 14th day of infection, becoming negative between the 45th and the 54th day (Fig. 1). Cumulative mortality - Excluding the animals sacrificed during the course of the experiment, all the infected C. callosus as well as the intact controls survived the experimental course. Morphological aspects - The Table summarizes the data for interstitial matrix alterations as evaluated according to the degree of inflammation and fibrosis. An arbritary point scale was applied: (+) indicating mild focal or diffuse alterations; (++) moderate focal or diffuse and (+++) diffuse and severe alterations, as previously described (Andrade et al. 1991). Histopathologic study - Normal controls - Sections of the heart showed the normal structure of the myocardium with scarce interstitial matrix components. Skeletal muscle of normal calomys stained with hematoxylin and eosin showed the characteristic histologic pattern, with a distinct slender perimysial membrane. Picro Sirius staining for collagen showed fine strands of perimysial collagen deposits. Infected C. callosus - The degree of the inflammatory infiltration in the myocardium and skeletal muscle is presented in the Table. Heart - In the early infection the myocardium presented focal and diffuse mononuclear inflammatory infiltrates with a predominance of macrophages and lymphocytes (Fig. 2 A); proliferation of fibroblasts with interstitial matrix deposits was also present (Fig. 2B). In sections stained with Picro-Sirius method, collagen was identified by positive bi-refringence to polarized light (Fig. 3A). Parasites were scarce or even absent in the heart. At the 40th day, small foci of destroyed cardiac cells with focal inflammatory infiltrate were present. (Fig. 2F, G). Picro Sirius staining for collagen showed fragmentation and a progressive decrease of collagen until the 90th day, with remnants of perivascular deposits and thin interstitial strands of collagen (Fig. 3B). Skeletal muscle - In the early infection, focal lesions of arteritis and peri-arteritis were present as well as an intense inflammatory lesions corresponding to focal destruction of muscle fibers and infiltration with macrophages, lymphocytes and fibroblasts (Fig. 2C). Large collections of well-preserved amastigotes of T. cruzi were present in skeletal muscles (Fig. 2D). Sections stained with Picro-Sirius showed the presence of strands of collagen with characteristic red staining and intense bi-refringence in the inflammatory foci, when examined with polarized light (Fig. 3C). At the 40th day of infection the inflammatory lesions became less intense and were reduced to moderate interstitial infiltrates with scarce matrix deposits. From 60th through the 90th day, the inflammatory infiltrates in skeletal muscles were focal and scarce, with areas of substitution of destroyed muscle fibers by adipose tissue (Fig. 2E, H). Fine strands of interstitial collagen appeared in sections stained with Picro-Sirius (Fig. 3D) . Immunolabelling of the matritial components of the skeletal muscles and myocardium Normal controls - In the skeletal muscle, the perimysial septa around groups of muscle fibers showed small deposits of Types I, III and IV collagens (Fig. 4A, D, G). In the sections immunolabelled for laminin and fibronectin, small deposits were seen in the perimysium and in small endomysial capillaries (Fig. 4J, N). The heart presented scarce interstitial matrix components with fine fibrilar deposits of collagens types I and III; collagen type IV and laminin apeared as fine deposits in the basal membrane of myocytes. Fibronectin was not detected.
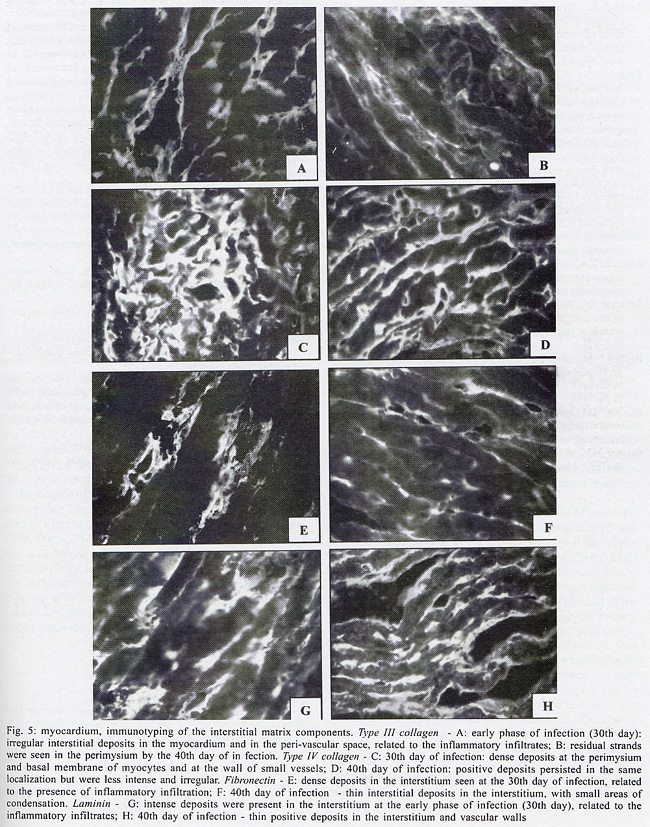
Infected C. callosus Skeletal muscle - Positive deposits immunolabelled for collagens types I, III and IV, laminin and fibronectin were present and varied from the early to the late infection. In the early phase (30th day of infection) the deposits were more intense and correlated with the presence of intense inflammatory infiltrates. Types III and IV collagens predominate in the early phase (Fig. 4E, H). Type I collagen deposits were less prominent (Fig. 4B). A clear decrease of types I, III and IV was seen from the 40th to the 60th day and appeared as fine, irregular and fragmented deposits by the 90th day (Fig. 4C, F, I). Laminin and fibronectin predominates by the 30th day (Fig. 4L, O) and also decreased at the same periods (Fig. 4M, P). Myocardium (Fig. 5A, H) - Type I collagen was seen as thin filaments in the inflammatory infiltrates and disappeared in the late phase of infection; deposits of collagens types III and IV were prominent in the early phase of infection (30th day) (Fig. 5A, C); they decreased from the 40th day (Fig. 5B, D), becoming residual or absent at the late phase (90th day). Fibronectin and laminin appeared during early infection (30th day) as irregular deposits in the interstitium, related with inflammatory foci (Fig. 5E, G) decreasing by the 40th day (Fig. 5F, H). At the 90th day, only fine and scarce deposits in the basal membrane were present. DISCUSSION In the present investigation, the immunolabeling of the interstitial matrix of the heart and skeletal muscles in C. callosus experimentally infected with T. cruzi demonstrated the participation of Types I, III and IV of collagens, laminin and fibronectin. A clear relationship between the regression of inflammation and the decrease in extracellular matrix deposits was detected. The reversibility of myocardial fibrosis has previously been described in murine chronic Chagas' myocarditis (Andrade et al. 1991), after specific treatment with anti-T. cruzi chemotherapy. The curative treatment induced regression of the inflammatory lesions and progressive changes in collagen, resulting in resorption of fibrosis (Andrade et al. 1991). In C. callosus the regression of the inflammatory infiltrate and collagen degradation occurred spontaneously, without interference of a specific treatment. It is not well understood which are the factors involved in this peculiar modulation. In both models, regression of parasitism and inflammation are of paramount importance for the regression of fibrosis. As suggested by Andrade (1994), the chronic inflammatory process in parasitic infections acts as potent promoter of matrix formation, and it is possible to demonstrate the degradation of excess matrix deposits, when this cause is removed and the inflammatory process subsides. In C. callosus infected with T. cruzi, the decrease of the fibrotic-inflammatory lesions occurred rapidly and corresponded to a decrease of parasite load, even with persistence of infection, as shown by the positive xenodiagnosis found in chronically infected calomys (Andrade et al. 1994). The evolution and regression of fibrosis in T. cruzi infection results from a peculiar host-parasite relationship, characterized by a delayed type hipersensitivity response in the host (Andrade et al. 1987, Ribeiro-dos-Santos et al. 1991, Dos Reis 1997), possibily followed by a down regulation of this mechanism (Andrade et al. 1994). Early deposits of collagen in the C. callosus model are probably related to an intense production of fibrogenic cytokines by activated macrophages as shown in mice (Zhang & Tarleton 1996). Collagen deposits are also regulated by factors determining an increase of fibrolysis of matrix deposits, such as the metalloproteinases (Zhang et al. 1999) and the tissue inhibitor metallopro-teinases (Eickelberg et al. 1997). Comparing the characteristics of the interstitial matrix in mice and callomys, it was seen that the predominance of types III and IV in the infected calomys is similar to that described in mice (Andrade et al. 1989), but differed in relation to the participation of type I collagen, demonstrated in calomys but not in mice. Several authors have emphasized the influence of the genetic type of collagen in the resorption of the fibrosis in the liver (Rojkind & Dunn 1979, Grimaud et al. 1980). However, Andrade and Grimaud (1986) and Andrade (1992), studying the regression of fibrosis in schistosomal granuloma have shown that both Types I and III collagens disappeared gradually and simultaneously. Results of the present study indicate that there is a clear correlation between the reversibility of fibrosis in C. callosus model and the early and spontaneous control of parasite multiplication in this model. In the infected calomys, the different components of the extracellular matrix were resorbed at the same period of infection, simultaneous to the regression of the inflammatory process, independent of collagen genetic types. ACKNOWLEDGEMENTS To Mr Brendan L Flannery (University of California, Berkeley, CA, USA) for the English language review. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02135f2.jpg] [oc02135f5.jpg] [oc02135f4.jpg] [oc02135f3.jpg] [oc02135f1.jpg] [oc02135t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}