 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(5) July 2002, pp. 717-724 Ultrastructural Alterations in Adult Schistosoma mansoni Caused by Artemether Xiao Shuhua, Shen Binggui, Jürg Utzinger*/**, Jacques Chollet*, Marcel Tanner*/+ Institute of Parasitic Diseases, Chinese Academy of Preventive Medicine, Shanghai, China *Swiss Tropical Institute, P.O. Box, CH-4002 Basel, Switzerland **Office of Population Research, Princeton University, Princeton, NJ, USA, +Corresponding author. Fax: +41-61-271.7951. E-mail: marcel.tanner@unibas.ch This investigation received financial support from the Ninth Five-Year Key Research Program of China, the UNDP/World Bank/WHO Special Program for Research and Training in Tropical Diseases (TDR) and the Swiss Tropical Institute. JU received financial support from the Swiss National Science Foundation and the Centre for Health and Wellbeing at Princeton University. Received 22 October
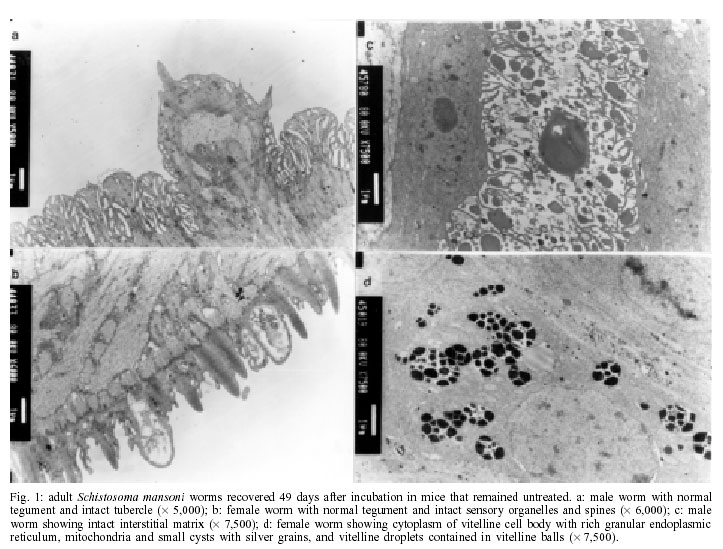
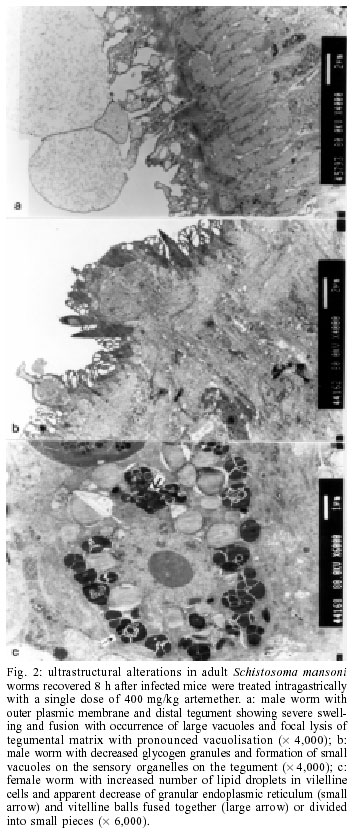
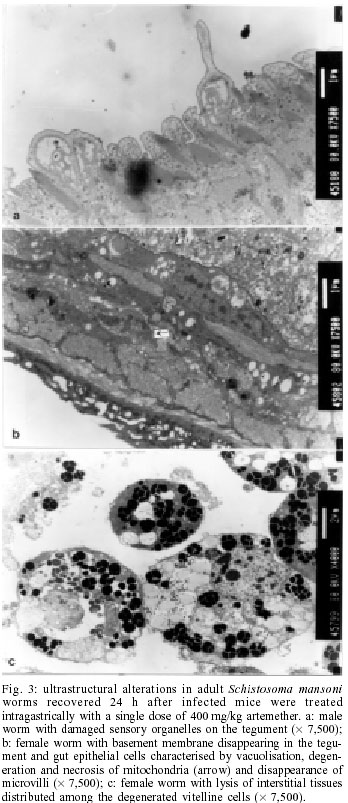
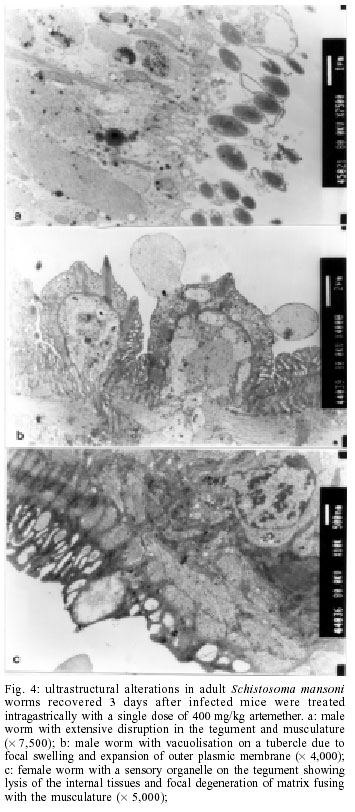
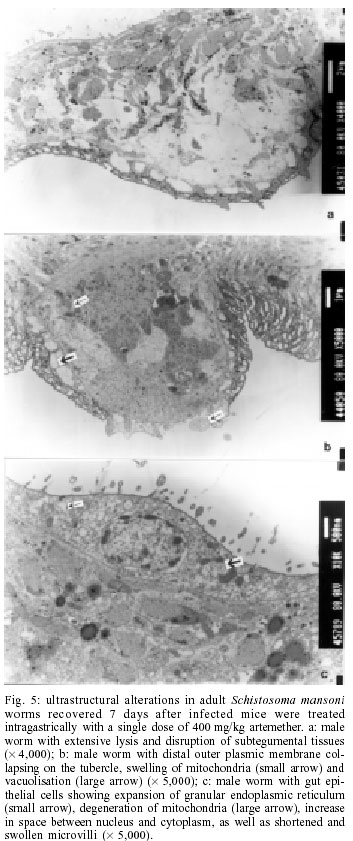
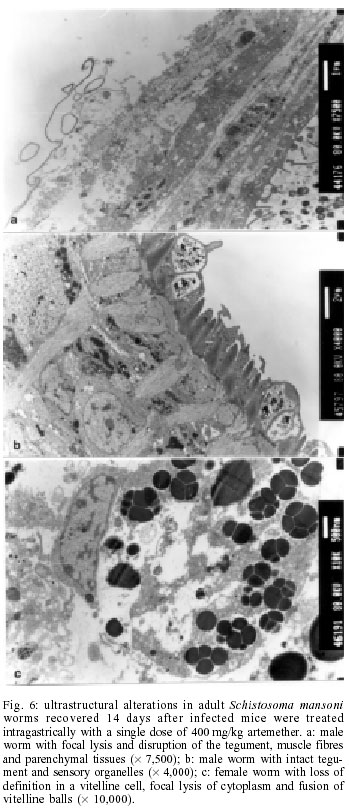
2001 Code Number: oc02137 Progress has been made over the last decade with the development and clinical use of artemether as an agent against major human schistosome parasites. The tegument has been identified as a key target of artemether, implying detailed studies on ultrastructural damage induced by this compound. We performed a temporal examination, employing a transmission electron microscope to assess the pattern and extent of ultrastructural alterations in adult Schistosoma mansoni harboured in mice treated with a single dose of 400 mg/kg artemether. Eight hours post-treatment, damage to the tegument and subtegumental structures was seen. Tegumental alterations reached a peak 3 days after treatment and were characterized by swelling, fusion of distal cytoplasma, focal lysis of the tegumental matrix and vacuolisation. Tubercles and sensory organelles frequently degenerated or collapsed. Typical features of subtegumental alterations, including muscle fibres, syncytium and parenchyma tissues, were focal or extensive lysis, vacuolisation and degeneration of mitochondria. Severe alterations were also observed in gut epithelial cells and vitelline cells of female worms. Our findings of artemether-induced ultrastructural alterations in adult S. mansoni confirm previous results obtained with juvenile S. mansoni and S. japonicum of different ages. Key words: schistosomiasis - Schistosoma mansoni - artemether - transmission electron microscopy - ultrastructural studies - adult worms Artemisinin and related derivatives are effective antimalarial drugs clearing parasitaemia and disease-related symptoms more promptly than any other currently available antimalarials (McIntosh & Olliaro 2000). The active principle stems from the leaves of Artemisia annua, a herbal plant used for centuries in Chinese traditional medicine (Klayman 1985). Several million malaria patients in Southeast Asia have been treated with artemisinins over the last two decades (Price et al. 1999). These drugs, characterised by very short terminal half life times, are now also becoming available in Africa. Combination therapies with longer lasting antimalarials are being advocated as a strategy to prolong the length of time artemisinins will remain effective (White 1999, White et al. 1999). In 1982, Chinese parasitologists discovered that administration of artemether (methyl ether derivative of dihydroartemisinin) to mice infected with Schistosoma japonicum resulted in significant reductions of the schistosome worm burden (Le et al. 1982). Despite the importance of this finding, it was recognized only recently by the international research community, alongside a World Bank project, placing renewed efforts on schistosomiasis control in China (Yuan et al. 2000). There is now compelling evidence from laboratory investigations that S. japonicum, S. mansoni and S. haematobium are susceptible to artemether (Xiao et al. 1995, 2000a, b). The highest susceptibility is confined to the larval migratory stages. However, artemether also shows activity against adult schistosomes, characterized by hepatic shift, degeneration of reproductive organs and hence inhibiting egg formation, and decrease of worm body size, which eventually leads to death (Wu et al. 1983, Xiao & Catto 1989, You et al. 1992, Xiao et al. 1995). Randomized controlled clinical trials with artemether orally administered once every 2-3 weeks significantly reduced the incidence and intensity of patent infections (Utzinger et al. 2000, Xiao et al. 2000c). We have recently discussed how these findings could be translated into medical use in different epidemiological settings (Utzinger et al. 2001a, b). Scanning electron microscopic observations in S. japonicum, S. mansoni and S. haematobium of different ages recovered from host animals treated with artemether revealed that the tegument is a key target of artemether. Tegumental alterations included swelling, fusion of tegumental ridges, vacuolisation, peeling, erosion and sometimes collapse of the tegument (Xiao et al. 1996a, 2000d, e, 2001a). Transmission electron microscopic studies in juvenile and adult S. japonicum, and juvenile S. mansoni further showed that, apart from the tegument, artemether also induces ultrastructural changes in subtegumental structures (Xiao et al. 1996b, 2002). In the absence of comparable data for adult S. mansoni (Utzinger et al. 2001b), we recovered schistosomes from mice 8 h to 14 days after the administration of artemether and assessed ultrastructural alterations by means of transmission electron microscopy. MATERIALS AND METHODS Ethical clearance, host animals, drugs and parasites - We obtained ethical clearance for our animal studies from the local government, which is in accordance with national regulations of Switzerland. The experiments were conduced in the laboratories of the Swiss Tropical Institute and followed standardized procedures described elsewhere (Xiao et al. 2000e). Twelve female MORO strain mice, weighing approximately 20 g, came from the Biological Research Laboratories LTD (Füllinsdorf, Switzerland). Mice were fed with Rodent Blox from Eberle NAFAG (Gossau, Switzerland; article no. 850.25W10), and were given water ad libitum. Artemether was the gift of the Kunming Pharmaceutical Corporation (Kunming, China; lot no. 97080) with a documented purity of 99.6%. Artemether was suspended in 7% Tween-80 and 3% ethanol and had a final drug concentration of 40 g/l. The volume of artemether suspension given to mice was 10 ml/kg. S. mansoni cercariae (Liberian strain) were collected from infected Biomphalaria glabrata snails, after exposure to artificial light for at least 4 h. Infection, artemether treatment and dissection - Mice were infected subcutaneously with 120 S. mansoni cercariae each. Seven weeks post-infection, 10 of 12 mice were treated intragastrically with a single dose of 400 mg/kg artemether. Treated mice were sacrificed, two at a time, by blood-letting at 8 h, 1, 3, 7 or 14 days post-treatment. Those two mice that remained untreated were also killed 7 weeks post-infection. Adult schistosomes were recovered by a perfusion technique, using ice-cold Hanks' balanced salt solution (HBSS) (Yolles et al. 1947). The schistosomes collected from the group of untreated mice served as control. Transmission electron microscopy - Recovered schistosomes were rinsed 3 times with ice-cold HBSS. The specimens were fixed in 2.5% glutaraldehyde-phosphate buffer (0.2 mol/l, pH 7.4) and kept at 4°C. Post-fixation was performed with osmium tetraoxide in the same buffer. Specimens were dehydrated in graded ethanol before embedding in Spur resin. Ultrathin sections of the schistosomes were stained with uranyl acetate and lead citrate. Ultrastructural features of the schistosome specimens were then examined, using a JEM-100B transmission electron microscope. RESULTS The ultrastructural features of the tegument and subtegumental structures of adult S. mansoni, recovered from untreated mice, are presented in Figs 1a-d. These control experiments revealed no apparent damage, as ultrastructural features were similar to those described in previous studies (Smith et al. 1969, Hockley 1973, Erasmus 1975). Administration of 400 mg/kg artemether to mice infected with 49-day-old S. mansoni resulted in ultrastructural alterations, which were apparent 8 h after recovery of the schistosomes. The first changes in the tegumental integrity of male worms were focal swelling of the distal cytoplasmic processes, accompanied by an increase in their width and loosening of the matrix. Alongside these focal alterations, there was formation of many small vacuoles. In some sections of the damaged tegument, the distal cytoplasmic processes expanded and fused together to form large vacuoles (Fig. 2a). The major alterations observed in female schistosomes included vacuolisation, focal lysis of the tegumental matrix and underlying muscle fibres. In the syncytium we observed emergence of myeline-like small bodies, disappearance of granular endoplasmic reticulum, and loss of definition of the tegument. The sensory organelles at the surface of adult worms showed a decrease in glycogen granules, lysis of mitochondria and formation of small vacuoles because of focal disruption of this portion of the tegument (Fig. 2b). In the tubercles, the distal tegumental matrix and the spines usually lysed. This process was accompanied by vacuolisation and a decrease in glycogen grains inside the tubercles. Meanwhile, expansion of granular endoplasmic reticulum, swelling of mitochondria, appearance of membrane-like inclusion bodies, formation of vacuoles in cytoplasm and shortening of microvilli was seen in gut epithelial cells. In vitelline cells, decrease or disappearance of granular endoplasmic reticulum and mitochondria, increase in lipid droplets and focal or complete fusion of vitelline balls in vitelline drops were the main alterations (Fig. 2c). In male worms the external appearance of the tegument, including tubercles and sensory organelles, as well as the tegumental matrix showed extensive lysis and indistinction 24 h post-treatment (Fig. 3a). At some locations, the basal membrane of the tegument disappeared, or the outer plasmic membrane expanded to form vacuoles. Owing to the severe swelling of the tegument, some spines were completely surrounded by the tegument. Focal or extensive lysis of muscle fibres and parenchymal tissues was usually seen. In syncytium the chromosomes of the nuclei increased, while the granular endoplasmic reticulum and mitochondria disappeared. Tegumental lesions in female worms were similar to those observed in male specimens, but there was also appearance of vacuoles of varying sizes contained in residual bodies of the cytoplasm of gut epithelial cells. Other lesions included degeneration and necrosis of mitochondria and disappearance of microvilli (Fig. 3b). In vitelline cells, the interstitial tissues usually lysed, while the nuclei of some cell bodies disrupted or disappeared (Fig. 3c). Many lipid droplets, vacuoles, membrane-like inclusion bodies and myelin-like structures appeared in the cytoplasm of these cells. The small balls in the droplets fused together or divided into numerous fragments, while the nucleated membranes expanded or collapsed. Three days following the administration of artemether, more pronounced alterations in the tegument and the musculature were observed in male schistosomes (Fig. 4a). Damage was also apparent in sensory organelles with some of them showing fewer mitochondria and increased density of glycogen grains. In the tubercles, there was lysis of tegumental matrix and spines and decrease in glycogen grains. Some tubercles showed extensive lysis in their internal structures, accompanied by formation of different size of vacuoles filled with residual bodies. There were other tubercles where the focal outer cytoplasmic membrane was damaged, resulting in the protrusion of the internal tegumental matrix and formation of vacuoles (Fig. 4b). In female worms, the sensory organelles and the tegumental matrix exhibited extensive lysis. The musculature showed extensive lysis, which resulted in further fusing with the tegument (Fig. 4c). The main lesion of the syncytium included disappearance of granular endoplasmic reticulum, degeneration of mitochondria, and lysis of cytoplasm or emergence of myelin-like structures. In gut epithelial cells expansion of granular endoplasmic reticulum, degeneration of mitochondria and loose distribution of microvilli were seen (Fig. 4d). The main alterations in vitelline cells were similar to those 24 h post-treatment, but the vitelline balls in the vitelline droplets fused together or collapsed (Fig. 4e). Seven days post-treatment, some male and female worms still showed severe damage and extensive lysis in the tegument and in the underlying muscle fibres and parenchymal tissues (Fig. 5a). The sensory organelles and tubercles of these worms still exhibited alterations (Fig. 5b). The extent and degree of expansion of granular endoplasmic reticulum and degeneration of mitochondria in gut epithelial cells were similar as 3 days post-treatment. In some gut epithelial cells the microvilli showed reduction in number, shortening in length and expansion at the end of the microvilli (Fig. 5c). In some ultrathin sections, the external appearance of the tegument was normal, suggesting that at least partial tegumental repair occurred. Fourteen days after artemether administration, there were some male and female worms where the most severely damaged parts of the tegument were completely detached from the worm surface (Fig. 6a). However, in other specimens, the damage was only focal with partial lysis observed on the tegument, musculature and parenchymal tissues. Intact tegument and muscle fibres were seen in some worms, as well as apparently unchanged sensory organelles and tubercles (Fig. 6b). Meanwhile, the gut epithelial cells still showed apparent lesions, including expansion of granular endoplasmic reticulum, partial degeneration or disappearance of mitochondria and emergence of numerous membrane-like vacuoles. However, more and longer microvilli were seen. The vitelline cells still remained severely damaged, as there was extensive lysis in the interstitial tissues among the cells. Some vitelline cells collapsed, which resulted in the release of vitelline droplets and emergence of myelin-like structures in the cytoplasm (Fig. 6c). DISCUSSION The tegument of schistosomes and other trematode species is a living, anucleate, and cytoplasmic structure. It is connected with underlying nucleated cells that are located beneath the circular and longitudinal musculature (Hockley 1973). The role the tegument plays at the interface of the parasite and the host has been investigated in numerous studies. It has secretory functions, is involved in nutrient absorption, shields schistosomes from the immune response by the infected host, and is a key target for antischistosomal drugs (Pappas & Read 1975, Capron et al. 1987, Pearce & Sher 1987). Therefore, detailed ultrastructural observations of the tegument and subtegumental structures following the administration of antischistosomals will further enhance our understanding of the host-parasite relationship (Hockley 1973). Alterations in the ultrastructure of S. mansoni worms have been documented for various antischistosomal drugs: Astiban (Erasmus 1975, Otubanjo 1981), hycanthone (Hillman et al. 1977), niridazole (Popiel & Erasmus 1981), oxamniquine (Popiel & Erasmus 1984, Magalhães Filho et al. 1987, Fallon et al. 1996) and praziquantel (Becker et al. 1980, Mehlhorn et al. 1981, Shaw & Erasmus 1983, 1987, 1988, Fallon et al. 1996). In common, all drugs induced damage to the tegument, subtegumental structures and the reproductive system of the worms. However, the onset, evolution and the precise pattern of these ultrastructural alterations dependent upon the particular drug used. Artemether is a new type of antischistosomal drug, but the exact mechanism of action by which this compound affects schistosomes has yet to be fully elucidated (Xiao et al. 2000c, Utzinger et al. 2001a, b). Artemether displays a considerably slower action than praziquantel (Cioli et al. 1995, Xiao et al. 2000f), but more research is required to explain this difference. When adult S. mansoni were recovered from mice 8 h after a single dose of 400 mg/kg artemether, ultrastructural damage was seen at some parts of the worm surface. The first and most common feature was vacuolisation of the tegument, which increased in severity to reach a peak 3 days post-treatment. For comparison, vacuolisation following the administration of praziquantel was observed as early as 5 min post-treatment (Becker et al. 1980). Artemether-induced vacuolisation of the tegument was accompanied by swelling of distal cytoplasmic processes and focal or even extensive lysis of the tegumental matrix. Tubercles and sensory organelles on the tegument showed various degrees of damage, and some tubercles completely collapsed. Subtegumental structures, including musculature, syncytium and parenchymal tissues, gut epithelial cells and vitelline cells, also showed extensive damage. However, in some schistosomes, partial tegumental repair occurred as there was less pronounced damage and intact appearing tubercles and sensory organelles 7 and particularly 14 days after cessation of artemether administration. Our findings of artemether-induced damage in the tegument of adult S. mansoni worms correspond with previous scanning electron microscopic observations (Xiao et al. 2000e). Recently, we have performed a temporal examination of ultrastructural alterations in 21-day-old juvenile S. mansoni. Infected mice were treated with artemether at the same dose and route of administration as in the experiments presented here and schistosomula were recovered between 8 h and 14 days post-treatment (Xiao et al. 2002). The occurrence, pattern and intensity of ultrastructural changes in juvenile S. mansoni were similar as described in the present work. At first site, these observations are quite surprising, because the juvenile stages of S. mansoni were found to be significantly more susceptible to artemether than the adult worms (Xiao & Catto 1989, Xiao et al. 2000a). Stage-specific artemether susceptibilities with highest activity in the larval developmental stages were also found for S. japonicum (Xiao et al. 1995) and S. haematobium (Xiao et al. 2000b). The determinants explaining the facts that ultrastructural alterations in juvenile and adult schistosome worms are comparable, but that artemether susceptibility is stage-specific remain to be established. Three possibilities are offered for discussion. First, the tegumental damage might lead to disappearance of the immunological `disguise' of the worm. This is believed to be of prime importance in causing the death of the worms (Shaw & Erasmus 1987). It is possible that the disruption of the tegument caused by artemether was not extensive enough _ at least at the dose levels used in our experiments _ resulting in lower worm reduction rates. Second, the process of tegumental repair can occur relatively rapid (Popiel et al. 1985). Therefore, it might be suggested that focal damage induced by an antischistosomal drug in the tegument of either juvenile or adult S. mansoni might be repaired effectively over the course of 7-14 days after cessation of the drug. In case of severe tegumental damage the host immune response might impact the repair process. Thus, tegumental repair in the surviving worms suffering from severe damage might be slower than in specimens with less damage. In two previous studies, employing a subcurative dose of praziquantel or amoscanate, the recovery process for repairing damaged tegument in adult S. mansoni worms was 65 and 102 days, respectively (Voge & Bueding 1980, Shaw & Erasmus 1987). Although we made no attempt to determine the effect of artemether over similarly long periods post-treatment, we observed partial repair of the tegument and subtegumental structures beginning at 7 days post-treatment. Third, recent in vitro experiments suggested that artemether interacts with haemin. It is believed that artemether is activated by haematin, cleaving its endoperoxide bridge to generate free radicals (Meshnick et al. 1996). This might be the ultimate cause of schistosome death (Xiao et al. 2001b). In view of higher percent worm reductions in juveniles compared to adults, under the same dose level of artemether, it is reasonable to suggest that the anti-oxidant system in adult worms is stronger than in immature worms (Nare et al. 1990). Artemether also showed severe damage to the gut epithelial cells and vitelline cells, which might further impact the physiological functions of the intestine, and the production of eggs by female worms. In vitro studies with schistosomes kept in media containing artemether together with haemin showed intestines with marked changes, including extension, distortion, and depigmentation (Xiao & Catto 1989, Xiao et al. 2001b). These changes are clear indications of disturbance of the digestion process. On the other hand, the vitelline gland of female worms occupied about one-half of the posterior portions of the worm body, providing vitelline cells that are necessary for the development of eggs. The damage of these cells usually impacts the formation and production of eggs. However, in surviving female worms egg production gradually recovered about two weeks post-treatment. In conclusion, a single dose of artemether induces ultrastructural alterations to the tegument and subtegu-mental structures of adult S. mansoni. These findings confirm previous transmission electron microscopic observations with juvenile S. mansoni and S. japonicum of different ages. Similar ultrastructural examinations will now be undertaken for S. haematobium to finally present the full picture of artemether-induced damage in the three major human schistosome parasites. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02137f1.jpg] [oc02137f6.jpg] [oc02137f4.jpg] [oc02137f5.jpg] [oc02137f2.jpg] [oc02137f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}