 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
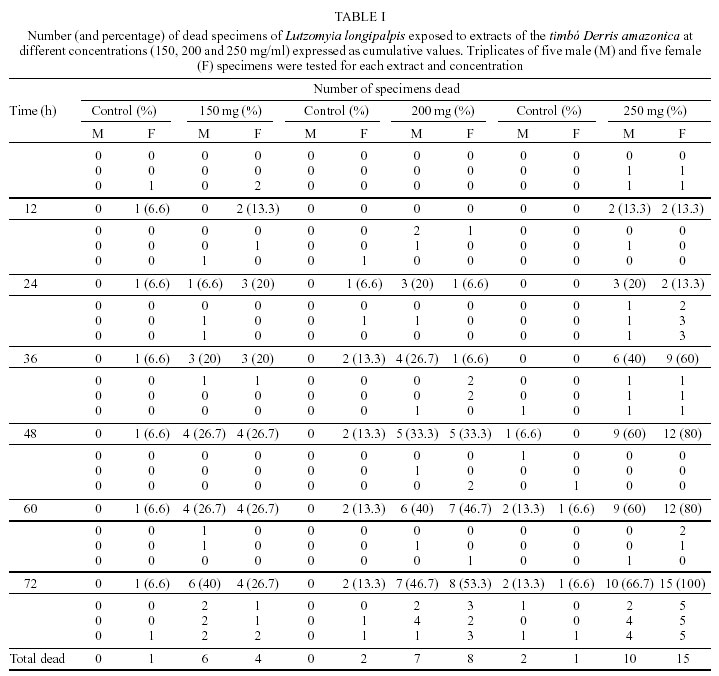
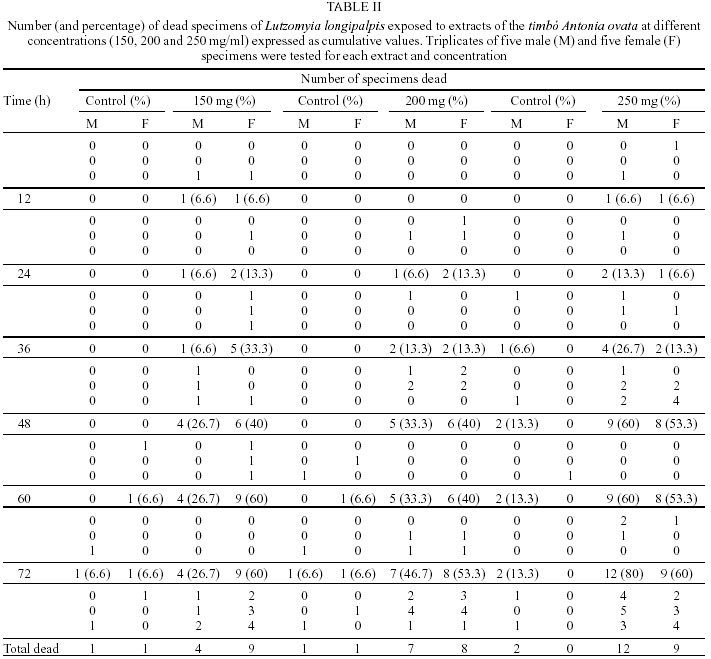
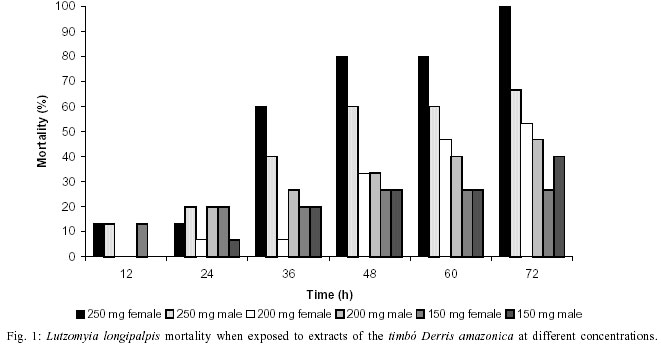
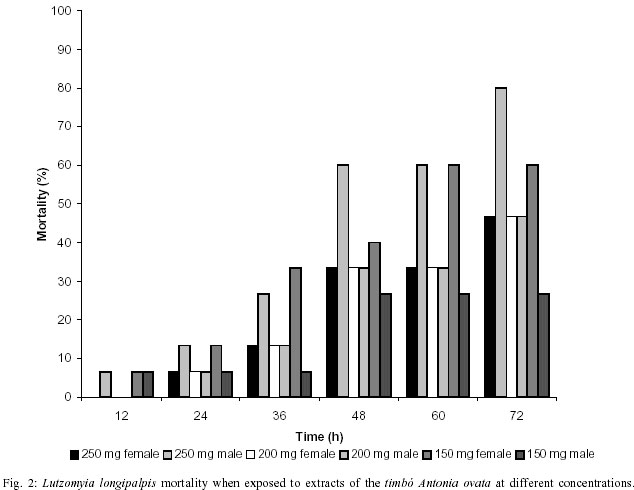
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(5) July 2002, pp. 737-742 Preliminary Assays Indicate that Antonia ovata (Loganiaceae) and Derris amazonica (Papilionaceae), Ichthyotoxic Plants Used for Fishing in Roraima, Brazil, Have an Insecticide Effect on Lutzomyia longipalpis (Diptera: Psychodidae: Phlebotominae) José Francisco Luitgards-Moura, Eloy Guillermo Castellón Bermudez*, Arnaldo Felisberto Imbiriba da Rocha**, Pantelis Tsouris, Maria Goreti Rosa-Freitas/***/+ Núcleo Avançado de Vetores do Instituto Oswaldo Cruz-Fiocruz, Centro de Ciências Biológicas e da Saúde, Universidade Federal de Roraima, BR 174 s/nº, 69310-270 Boa Vista, RR, Brasil *Instituto Nacional de Pesquisas da Amazônia, Manaus, AM, Brasil **Departamento de Química, Universidade do Amazonas, Manaus, AM, Brasil ***Departamento de Entomologia, Instituto Oswaldo Cruz-Fiocruz, Rio de Janeiro, RJ, Brasil, +Corresponding author. Fax: +55-21-2573.4468. E-mail: goreti@hotmail.com This work was partially supported by Capes (MSc scholarship to JFLM) and by the Government of Roraima. Received 31 August 2001 Code Number: oc02140 Laboratory-reared Lutzomyia longipalpis (Lutz and Neiva 1912) was tested with extracts of two ichthyotoxic plants, known as timbós, used as fishing poison in the Amazon. Phlebotomines, L. longipalpis, and plants, Antonia ovata and Derris amazonica, were collected in the Raposa-Serra do Sol Indian Reserve, a focus of visceral leishmaniasis in the State of Roraima, Brazil. Extracts were prepared from dried leaves of A. ovata and roots of D. amazonica that were percolated in water, filtered and dried out at 50°C. The solid extract obtained was diluted in water at 150, 200 and 250 mg/ml. The solution was blotted in filter paper placed at the bottom of cylindric glass tubes containing sand flies. For each plant extract and dilution, two series of triplicates with 5 male and 5 female specimens of L. longipalpis were used. Mortality was recorded every 2 h during 72 h of exposure. At 72 h the mortality was as high as 80% for extracts of A. ovata (LD50 = 233 mg/ ml), and 100% for D. amazonica (LD50 = 212 mg/ ml) whereas in the control groups maximum mortality never surpassed 13%. Preliminary assays indicated that A. ovata and D. amazonica displayed significant insecticide effect against L. longipalpis. Key words: ichthyotoxic extract - timbó - natural insecticides - Lutzomyia longipalpis - Derris amazonica - Antonia ovata - Roraima - Brazil Visceral leishmaniasis is considered to be a zooanthroponosis formerly restricted to rural and peri-urban areas in Brazil. Lately, visceral leishmaniasis has assumed an important role in public health due to the emergence of foci of the disease in urban areas (Marzochi & Marzochi 1994). Besides being the main visceral leishmaniasis vector in Brazil, Lutzomyia longipalpis can be a nuisance, as flies can bite at any time and at high densities (Quinnel & Dye 1994). Leishmaniasis transmission occurs mainly in the peridomicile although can also occur in the extradomicile (Lainson et al. 1985). Even though insecticide spraying is not routinely used for phlebotomine control, it has been used in situations where important peri- and intradomiciliary transmission prevailed (Marzochi & Marzochi 1994, Pereira & Fonseca 1994). The peridomestic vectors of Leishmania, Phlebotomus papatasi (in the Old World) and L. longipalpis and L. intermedia (in the New World) have been controlled by intradomiciliary insecticide spraying (Falcão et al. 1991, López-Antuñano 1992). Crude plant extracts and inorganic larvicides were largely used as natural insecticides before the organic laboratory synthesized insecticides became available in the 1940 decade (Balandrin et al. 1985, Raws 1986, WHO 1997). In fact, natural insecticides have been used for centuries to combat insect pests that compete for food and that affect public health (WHO 1997). Plant extracts such as pyrethrum, nicotine and rotenone were among the first compounds used to control insects of agricultural importance (Grainge & Ahmed 1988). Pyrethrins, a complex of esters extracted from flowers of Chrysanthemum cinerariefolium, are still used nowadays to enhance commercial preparations of household insecticides (Bell et al. 1990). Nicotine extracted from Nicotiana glauca and its nicotinoids derivatives are choice molecules for the manufacture of new insecticides. Rotenone and rotenoids are isoflavonoids occurring in several genera of tropical leguminosae plants such as Derris (Papilionaceae), Antonia (Loganiaceae) and Lonchocarpus (Fabaceae). There have been reports of rotenone-containing plants used by Indians to fish due to its ichthyotoxic effect, as early as the 17th century (Biet 1664 apud Moretti & Grenand 1982). Rotenoids have strong paralysis action on cold-blooded animals. For mammalians lethality tests resulted in LD50 as low as 25 mg/kg (Raws 1986). Rotenoids are also reported to have an insecticide effect against the tobacco budworm Heliothis virescens (Lepidoptera: Noctuidae; Yoshida & Toscano 1994). Rotenoid-yielding plants have being used for fishing based on their ichthyotoxic properties. Ichthyotoxicity is indeed an attribute distributed among a wide range of species. Mangrove Euphorbiaceae and Sapindaceae plants and umbellifer (Teixeira et al. 1984, Van Wagenen et al. 1988, Miles et al. 1989), dinoflagellates (Igarashi et al. 1999), cyanobacterium (Berman et al. 1999), blue-green and red tide algae (McCoy & Martin 1977, Igarashi et al. 1998), nudibranch and dictyoceratide sponge (McPhail et al. 1998) are among those reported as producing ichthyotoxins. Ichthyotoxicity, and insecticide activity for some of these species, make them of potential interest for use in agriculture, aquaculture and animal husbandry (Reinertaen et al. 1987, Johnson et al. 1988). Ichthyotoxic plants are known in Brazil under the generic denomination of timbó. The utilization of ichthyotoxic plants among native Amazonian Indians (Maiongong/ Ye'kuana, Wapixana, Macuxi and Sanumá ethnic groups; Farabee 1918, Smith 1939) has been done mainly as a fishing method. Timbós are in fact plants of many genera and species. Usually, roots of Derris sp. and Tephrosia sp. (Leguminosae) and leaves of A. ovata and Phyllanthus brasiliensis are broken and macerated with the aid of stones in wooden pestle and mortar. The macerated material is squeezed on partially blocked river streams, or lakes, giving off a white milky liquid with a characteristic smell. The previous downstream blockage use sticks of bamboo or guarumã (Ischnosiphon aruma) to increase effectiveness and retain the drugged fish. Interestingly, rotenone poisoned fish is edible without any risk of food contamination to humans (Costa et al. 1989). Others species of common use include D. utilis (timbó-macaquinho), D. urucu (timbó-urucu), Tephrosia toxicaria (timbó-de-caiena). D. amazonica is locally known as ailá and A. ovata as timbózinho (Angely 1985). A. ovata is widely distributed among fields and savannas of Brazil, but their use for fishing is restricted to the State of Roraima. Previous ichthyotoxic effect studies showed that the mechanism of insecticide action is not well understood and seems to be ubiquitous for insects. D. elliptica and D. uliginosa from Fiji showed insecticide activity (Mors et al. 1973). Purified rotenones from the roots of Derris and Lonchocarpus are used as insecticides (Sukamar et al. 1991). Triatomines exposed to extracts of D. urucu showed a mortality of 90-100% with a residual action up to 6 months (Klocke 1989). D. urucu and D. nicou also tested positive against lice and larvae of Musca domestica (Costa et al. 1989, 1997). In the literature there has been no report on the effects of D. amazonica and A. ovata on insects. The aim of this study was to verify the insecticide effect on L. longipalpis of A. ovata and D. amazonica, common ichthyotoxic plants used for fishing by indigenous people in Roraima. MATERIALS AND METHODS Collection areas - Plant and L. longipalpis phleboto-mine specimens were collected in the localities of Maloca da Malacacheta 02°40'11''N, 60°27'21''W (roots of D. amazonica Killip, Papilionaceae) and Maloca do Napoleão 03°55'04''N, 60°01'44''W (leaves of A. ovata Pohl, Loganiaceae), an endemic leishmaniasis area located in the Raposa-Serra do Sol Indian Reserve, Roraima, Brazil. Macuxi and Wapixana Indian ethnical groups mainly form local population. This endemic visceral leishmaniasis area is located in the Normandia municipality (03°52'52''N, 59°37'22''W, 100 m). The region is formed by savanna, with a tropical humid climate and maximum rainfall of 1,600 mm in June (Ministério das Minas e Energia 1975). L. longipalpis specimens were collected and bred in colony, maintained at 26 ± 2°C and 86% relative humidity (Luitgards-Moura et al. 2000). Plants and extraction - Specimens of timbó were collected together with individuals of the Macuxi and Wapixana communities. Plants were identified to species by Dr Aldaléa Sprada Tavares (Instituto Nacional de Pesquisas da Amazônia-Inpa, Manaus). Roots of D. amazonica and leaves of A. ovata were crushed in water to obtain extracts. Leaves and roots were allowed to dry in the shade, at room temperature (30° ± 5°C), crushed, weighted and submitted to extraction with hot water in a coffee maker (1 l capacity, Walita). Five liters of water were used to percolate 300 g of either roots or leaves (60 g/l). Infusion was filtered and water allowed to evaporate slowly by heating the extract at 50°C in a Pyrex glass dish on a hot plate (48 h average). Residue was weighed, scratched with spatula, transferred to a grail, crushed and placed in sealed jars in the refrigerator (16°C). Bioassays - L. longipalpis specimens were placed in glass vials with 5 x 8 cm filter papers blotted with different extract concentrations previously wet with distilled water and allowed to dry out overnight at room temperature. Filter papers were then placed at the bottom of the glass bottles and specimens transferred into them. A piece of gauze was used to close the bottle. To feed phlebotomines a piece of cotton, wet with 5% glucose solution, was placed on the gauze. Two triplicate series with 5 specimens of L. longipalpis were used for each plant extract (roots of D. amazonica and leaves of A. ovata) and dilution (150, 200 and 250 mg/ml). The first triplicate series contained 5 males in each vial and the second triplicate series contained 5 females in each vial. Therefore, 30 specimens were assayed for each plant extract and dilution. Negative controls used the same schedule. Males and females were not tested together. A total of 180 specimens was assayed (Tables I, II). Bottles containing males and females were placed in three separated plastic boxes according to dilution with a piece of wet filter paper at the bottom to keep moisture. Mean lethal dosage, designated as LD50, was determined at 12, 24, 36, 48 and 72 h of exposition. Acute toxicity was expressed by the necessary amount in mg of product per body weight to promote death of 50% of specimens. LD50 was calculated using the Polo program (Russel et al. 1977). RESULTS The LD50 was 233 mg/ml for A. ovata and 212 mg/ml for D. amazonica at 48 h of exposure. Apparently, L. longipalpis females were more affected by D. amazonica extracts, while males seem to be more affected by A. ovata. At 72 h, exposition to D. amazonica (250 mg/ml) resulted in 100% female mortality, opposed to 60% for A. ovata (Figs 1, 2). Male exposition to A. ovata 250 mg/ml extract resulted in 80% mortality, opposed to 66.7% for D. amazonica, at 72 h (Figs 1, 2). Curiously, A. ovata at 150 mg/ml resulted in higher mortality for females than did at higher concentrations, from 36 h onwards (Fig. 2). Maximum mortality in the control groups did not exceed 13.3% (Tables I, II). DISCUSSION Notwithstanding the advantageous eco-toxicological properties of natural insecticides and species selectivity and biodegradability, synthetic insecticides have almost completely replaced natural ones as the standard method of controlling detrimental insects, ticks and mites. Control programs for vector transmitted diseases are challenged by the increasing resistance of vectors to insecticides, the increasing costs of insecticides and the need for alternative control methods, including new insecticides (White 1991). Because of problems arisen with broad insecticide effects, soil and water contamination and resistance, the demand for natural and nonpersistent insecticides increased. Current biotechnology procedures allow the development of engineered plants and plant cell cultures that produce a wide array of insecticide products as part of their secondary metabolism in high amounts. Natural insecticide products originated from plants have been used since ancient times. Pyrethrum, saponin, nicotine and rotenone, all natural insecticides, have been largely used in pest control for medical and agricultural purposes (Coats 1994). Nowadays more than 2,000 plant species are catalogued as having insecticide properties, including the timbós (Heal et al. 1950, Farnsworth 1966, Sukamar et al. 1991). Insecticide action of timbó is related to the inhibition of the cellular respiration pathway (Aragão & Valle 1973). Insects display a decrease on oxygen consumption, on muscle contraction and finally paralysis when in contact with timbós (Aragão & Valle 1973). Toxic components belonging to the group of saponins and rotenones have been identified in timbós (Braz Filho et al. 1975). Saponins are phytoglycosides with emulsifying, toxic and hemolytic properties and also have the ability to complex cholesterol (Chacon 1973). Anti-fungicidal, anti-bacterial and abortive properties were also reported for saponins (Mahato et al. 1982, Moretti & Grenand 1982). Rotenone is believed to be the main toxic component in timbó species (Derris and Tephrosia, Chacon 1973, Xavier 1975). Separation of rotenoid active products from the raw material could potentialize insecticide effects observed in the extracts. Our preliminary results indicate that D. amazonica and A. ovata extracts have an insecticide effect on L. longipalpis males and females. Plant-derived substances are sources of potential new insecticides that may play a more prominent role in integrated pest management programs. Besides being candidates as biomolecules of choice for scale-up production, the use of timbós extracts could represent an economically viable way of vector control made by people living in these leishmaniasis endemic areas. D. amazonica and A. ovata are readily available and of common use by the community where visceral leishmaniasis is endemic in Roraima. Extracts are obtained through basic techniques and tools and, can be either sprayed on the walls or fumigated. New control insecticide substances can emerge from traditional popular knowledge bringing a sustainable vector control method in poor areas. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02140f2.jpg] [oc02140f1.jpg] [oc02140t1.jpg] [oc02140t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}