 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(6) September 2002, pp. 819-827 Cuticular Hydrocarbons of Chagas Disease Vectors in Mexico M Patricia Juárez/+, David A Carlson*, Paz María Salazar Schettino**, Sergio Mijailovsky, Gloria Rojas**
Instituto de Investigaciones Bioquímicas
de La Plata, Conicet-UNLP, Facultad de Ciencias Médicas, Calles 60 y
120, La Plata, 1900, Argentina *USDA, ARS, Medical and Veterinary Entomology
Research Laboratory, Gainesville, Fl, USA **Laboratorio de Biología de
Parásitos, Departamento de Microbiología y Parasitología,
Facultad de Medicina, UNAM, Mexico DF, Mexico This work was partially supported by grants from the AVINA Foundation, and the European Commission, and benefited from international collaboration through the ECLAT network and through Mexican research project, Conacyt 30871-N and WHO/TDR ID-970854 to PMSS. Received 17 December 2001 Code Number: oc02157
Capillary gas-liquid chromatography was used to analyse the cuticular hydrocarbons of three triatomine species, Triatoma dimidiata, T. barberi and Dipetalogaster maxima, domestic vectors of Chagas disease in Mexico. Mixtures of saturated hydrocarbons of straight and methyl-branched chains were characteristic of the three species, but quantitatively different. Major methylbranched components mostly corresponded to different saturated isomers of monomethyl, dimethyl and trimethyl branched hydrocarbons ranging from 29 to 39 carbon backbones. Sex-dependant, quantitative differences in certain hydrocarbons were apparent in T. dimidiata.
Key words: Triatoma - Dipetalogaster - cuticular hydrocarbons - gas chromatography - chemotaxonomy - Mexico Chagas disease (American trypanosomiasis), is widespread in the Americas, with the causative parasite, Trypanosoma cruzi, usually transmitted to humans in the faecal droppings of large blood-sucking insects of the subfamily Triatominae (Hemiptera, Reduviidae). In Mexico, over 30 species of Triatominae have been reported, mainly Triatoma species of the protracta and phyllosoma complex (Zeledón 1981, Zárate & Zárate 1985, Salazar Schettino et al. 1988, Schofield 2000). Most of them are actual or potential vectors of the disease, and Mexican authorities are currently carrying out surveys for Chagas disease vectors and implementing control trials in order to evaluate risk areas. As part of a larger study, the Latin American Network for Research on the Biology and Control of Triatominae is contributing to these efforts by evaluating the distribution and classification of domiciliated Triatominae using morphological, morphometric, molecular and biochemical techniques. Within this framework, we report here a study of the cuticular hydrocarbons of representative Mexican species of Triatominae, designed to evaluate the use of hydrocarbon profiles for species characterization and phylogenetic studies. MATERIALS AND METHODS
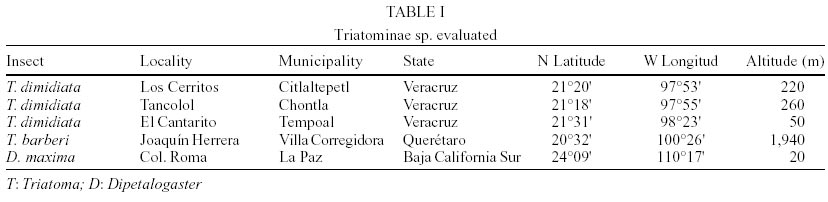
Insects - We analyzed hydrocarbons from wings of adult male and female individual specimens of T. dimidiata, collected from domestic and peridomestic locations from the municipalities of Tempoal, Citlaltepec and Chontla, in the State of Veracruz. The T. barberi specimens were collected from domestic habitats in the village of Joaquín Herrera, municipality of Villa Corregidora, State of Querétaro, and the Dipetalogaster maxima specimens were collected from houses in Colonia Roma, municipality of La Paz, State of Baja California Sur (Table I). Wings of each specimen were wrapped in aluminum foil and stored at room temperature prior to analysis. Hydrocarbon analysis - Cuticular hydrocarbons were extracted as previously described (Juárez & Blomquist 1993, Juárez et al. 2001). Wings from each specimen were washed with redistilled water to remove any water soluble contaminants, transferred to a glass vial with Teflon-lined caps, and submerged in redistilled hexane (6 ml/g) overnight, to extract total lipids. The solvent was transferred to another vial, reduced in volume under nitrogen, then hydrocarbons were separated from other components by adsorption chromatography performed on a mini-column of activated Biosil A (10 mm x 5 mm I.D.), eluting with redistilled hexane (6 ml/mg hydrocarbon). This final extract then was evaporated to an appropriate volume for gas chromatography. Capillary gas chromatographic (CGC) analysis was performed using a Hewlett-Packard (HP) Model 6890 gas chromatograph equipped with a cool on-column injector port and autoinjector system, fitted with a non-polar fused silica (0.2 µm) HP-5 capillary column (30 m x 0.32 mm I.D.), the carrier gas was H2 at a linear velocity of 40 cm/sec. The oven temperature was programmed from 60°C (hold time 2 min) to 180°C at 20°C/min, then 180°C to 310°C at 3°C/min (hold 10 min). The flame ionization detector (FID) was held at 320°C. A PC based data system, Turbochrom 3 (Perkin Elmer, CA, USA) was used for data recording and quantification. Injection of n-alkane standards of 22 to 42 carbons was similarly performed for estimation of Kovat Indices (KI) (Kovats 1965). Numbers showed close agreement with the KI's calculated for other triatomines (Juárez & Blomquist 1993, Juárez et al. 2001), and prediction of methyl branching pattern was done as proposed by Carlson et al. (1998). The nomenclature used to describe hydrocarbons was (Cn) to describe the total number of carbons in the corresponding hydrocarbon component; the location of methyl groups is indicated by (x- me) for monomethyl-alkanes, (x,x- dime) for dimethylalkanes and (x,x,x- trime) for trimethylalkanes. RESULTS
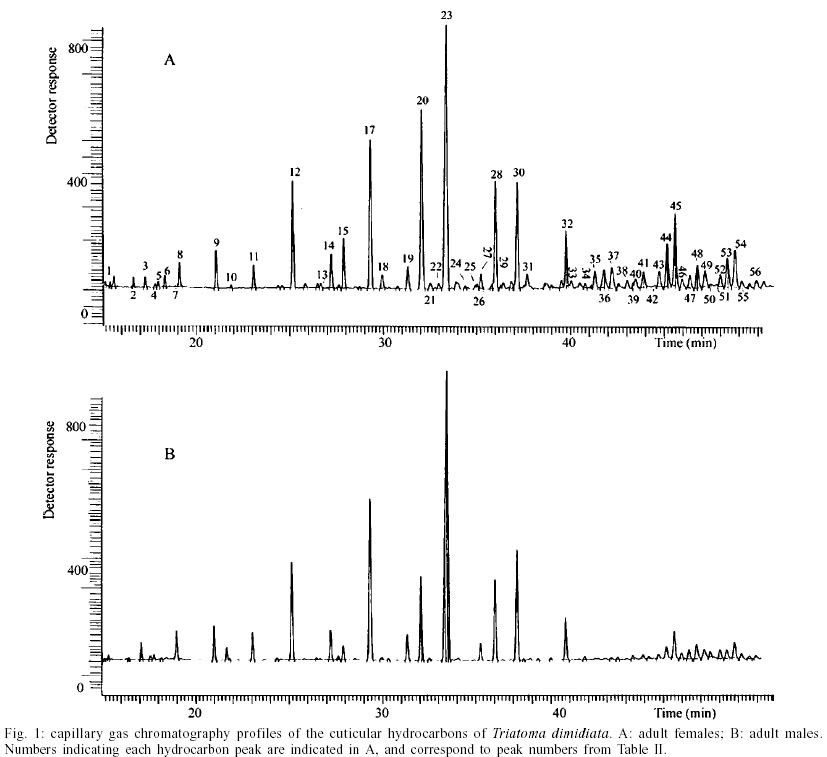
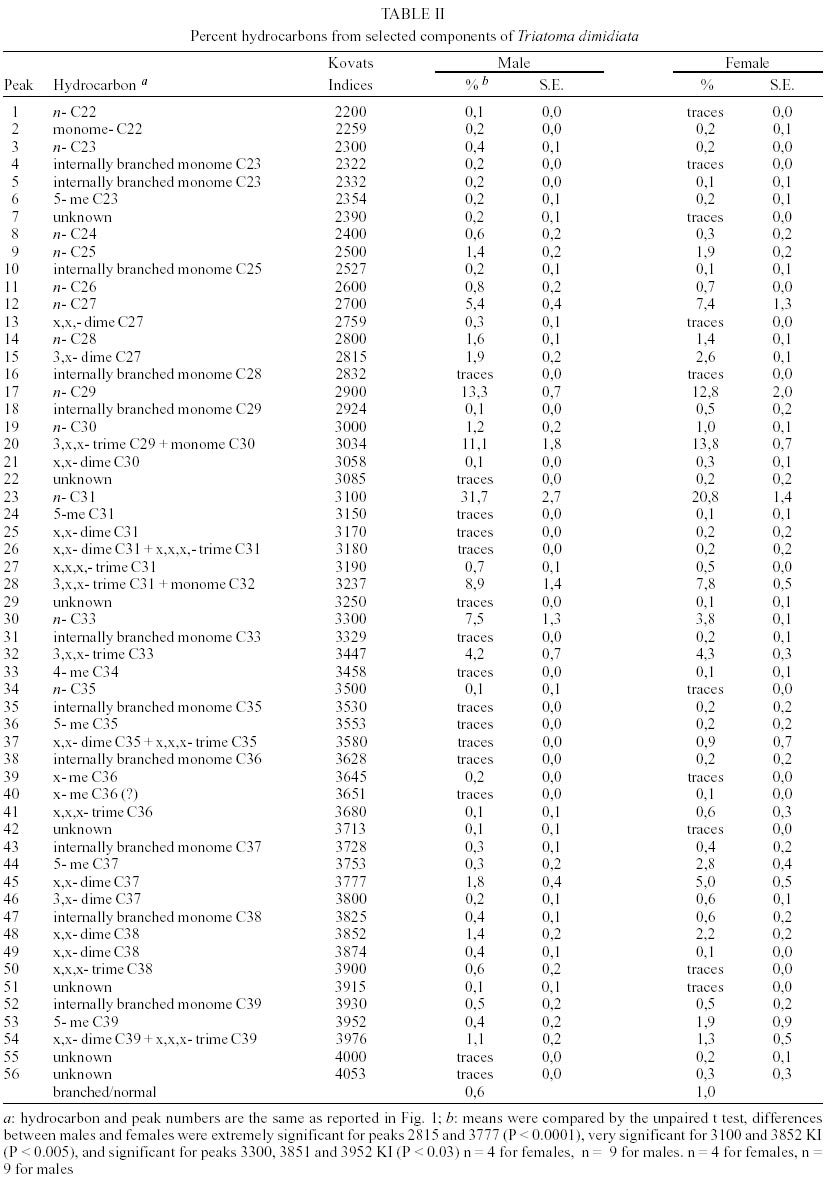
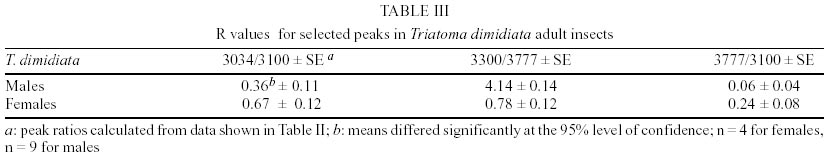
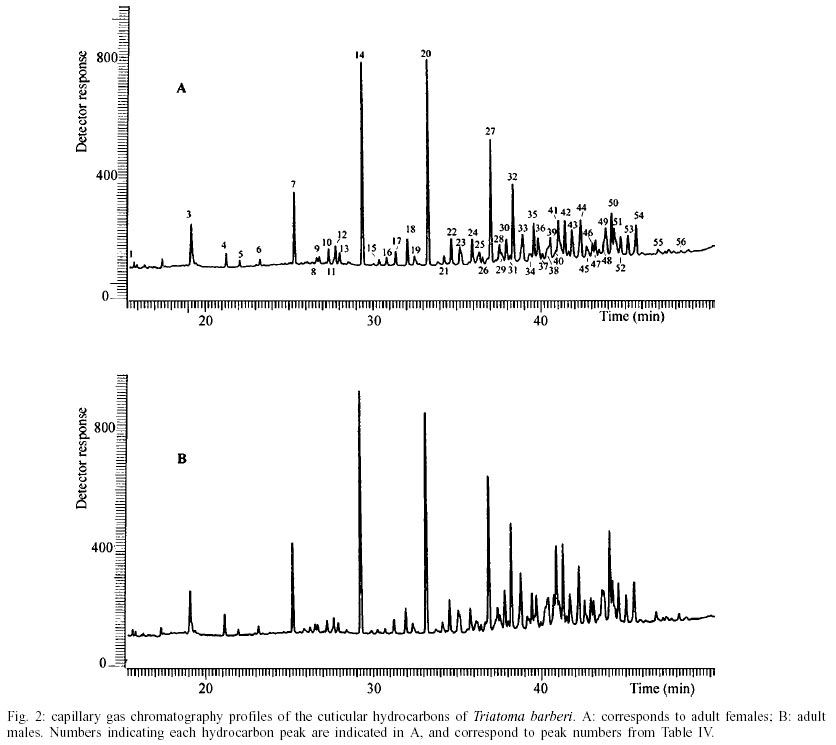
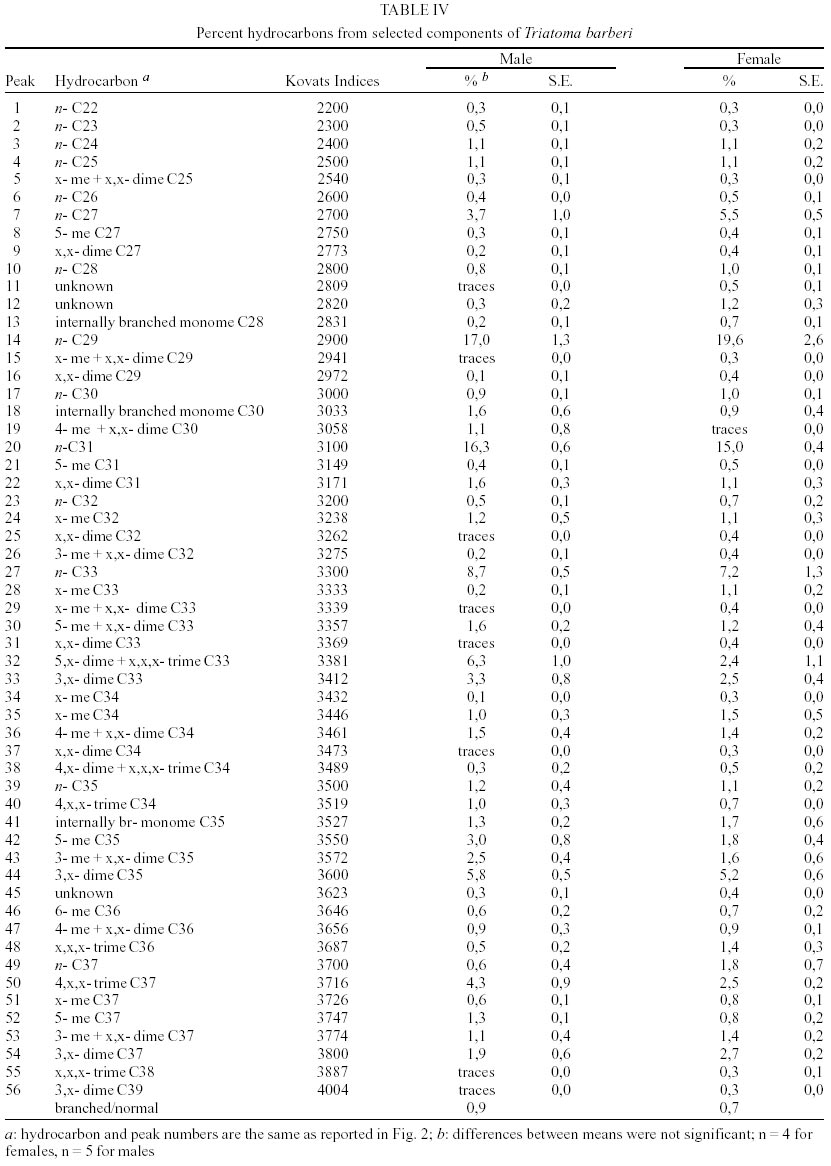
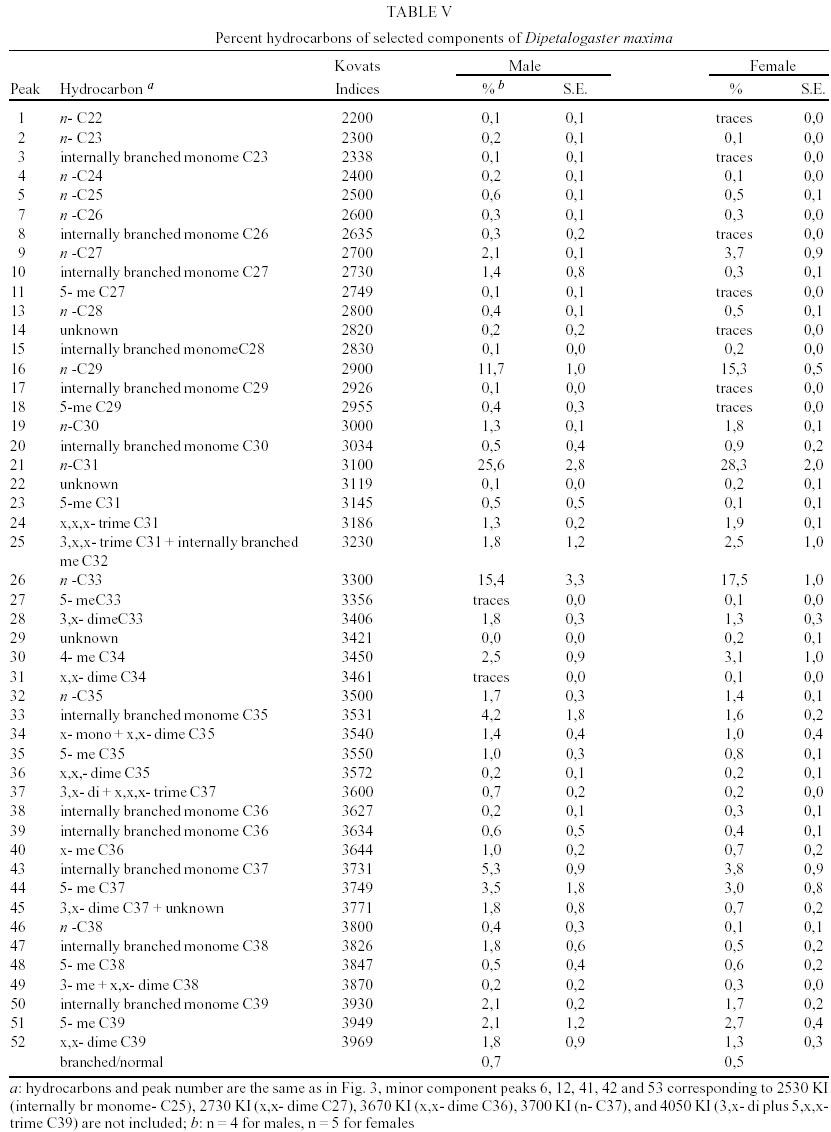
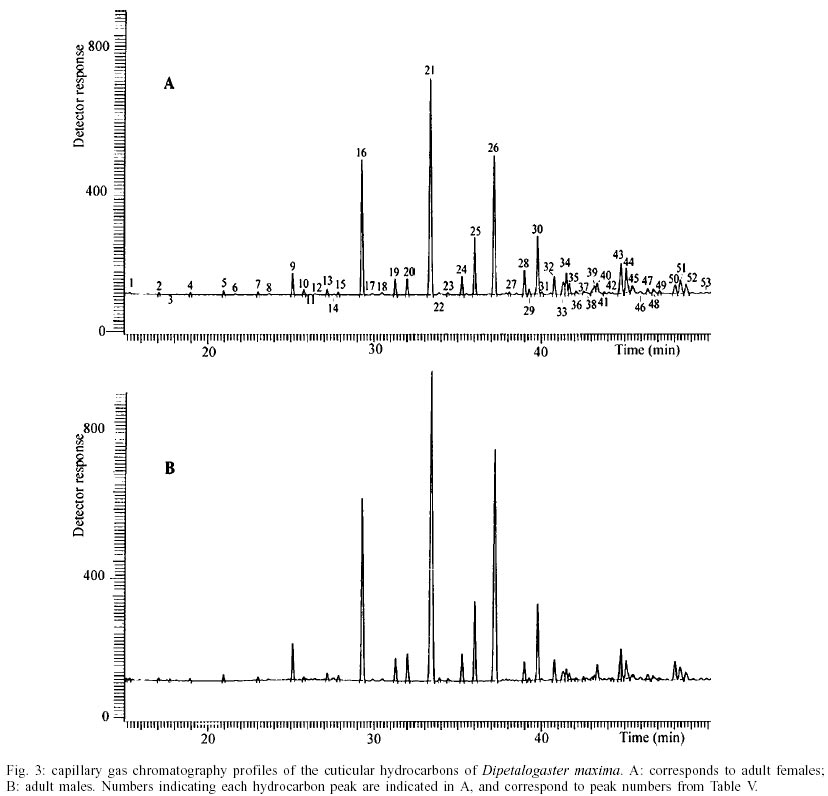
T. dimidiata - Typical gas chromatographic profiles of hydrocarbons extracted from individual male and female wings of T. dimidiata are shown in Fig. 1 A-B. The straight chain hydrocarbon components comprised saturated chains ranging from C22 up to 35 total carbons and accounted for 64.1% (males) and 50.4% (females) of the total wing hydrocarbon extract. Odd-chain components prevailed, with n- C31 accounting for 35.8% (males) and 20.9% (females) of the total hydrocarbon, followed by n- C29, n- C27 and n- C33; smaller amounts of even-numbered hydrocarbons, n- C22 through n- C30, were also detected (Table II). Among the methyl-branched alkanes, the major components consisted of a mixture of different isomers of terminally branched trimethyl odd chains together with internally branched monomethyl of even numbered chain, eluting at 3034 KI (3,x,x- trimethylC29 plus x- methyl C30) and 3237 KI (3,x,x- trimethyl C31 plus x- methyl C32), followed by 3,x,x- trimethyl C33 (3447 KI), 3,x- dimethyl C27 (2815 KI), and internally branched x,x- dimethyl C37 (3777 KI), x,x- dimethyl C38 (3852 KI) and at 3976 KI eluted a mixture of x,x- dimethyl and x,x,x- trimethyl C39. Sexual differences evaluated by the unpaired t test were very significant for the peak eluting at 3100 KI, the major hydrocarbon component n-C31, extremely significant (P < 0.005) for peaks of 3777 and 2815 KI (P < 0.0001) corresponding to x,x- dimethyl C37 and 3,x- dimethyl C27, respectively. Significant differences were found for peaks eluting at 3300, 3852 and 3952 KI (P = 0.02) (Table II). Four peaks were selected for peak ratios (R values) to evaluate sex-dependant, quantitative differences in hydrocarbon components, and the comparison of three R values separated T. dimidiata males and females at the 95% level of confidence (Table III). In addition, females had higher amounts of methyl-branched components than males with a branched to normal ratio of 0.98 for females and 0.56 for males. Peaks at 3150, 3170, and 3180 KI corresponding to mono-, di- and trimethyl C31 as well as mono-, di-, and trimethyl C35 eluting at 3530, 3553 and 3580 KI, and monomethyl C36 (3628 KI) were detecttable in females, but less apparent in males. T. barberi - Fig. 2A, B shows the gas chromatographic traces for adult males and females of T. barberi. The major n-alkanes are n-C29, n-C31, together with n-C33 and n-C27, plus minor amounts of straight chains from 22 to 37 carbons representing 57.9-52.5% respectively of the total hydrocarbons. Major methyl-branched components corresponded to mono-, di- and trimethyl derivatives of C33, C35 and C37 chains, with a branched to normal ratio of 0.90 for males and 0.73 for females. The KI values agreed well with one methyl group internally and subterminally located, dimethyl isomers with at least one methyl inserted internally, subterminal or terminally, and when three methyl groups were present, they were located at internal positions. Minor amounts of mono-, di- and trimethyl derivatives of C32, C34, C36 were observed as well as mono- and dimethyl- C25, C27, C29 and C31 chains (Table IV); differences between males and females were not significant. D. maxima - D. maxima hydrocarbon profiles showed a predominance of normal chains with a branched to normal ratio of 0.67 for males and 0.50 for females (Table V, Fig. 3), and no sexual dimorphism was evident. The major alkane was n- C31, followed by n- C29 and n- C33 which together accounted for 52.8% of total hydrocarbon for males and 61.1% for females. In the methylbranched fraction, three isomer series of 35, 37 and 39 atoms in the carbon backbone together with minor amounts of 4-me C34 and two series of 36 and 38 carbons were the prevailing branched structures. DISCUSSION
Analysis of insect cuticular hydrocarbons was shown to be useful for discriminating members of a number of insect complexes, among them the Anopheles gambiae complex (Carlson & Service 1979, Milligan et al. 1986), A. maculipennis complex (Phillips et al. 1988), and Glossina species (Carlson et al. 1993). Within the Triatominae, cuticular hydrocarbon analysis can help in differenciating species of the infestans complex (Juárez & Brenner 1985), and also provides phylogenetic markers for comparing the main genera, Triatoma, Rhodnius and Panstrongylus (Juárez et al. 2000). The hydrocarbon structures of the Mexican species T. pallidipennis and T. mazzotti, both included in the phyllosoma complex, were previously determined by CGC coupled to mass spectrometry (MS) (Juárez & Brenner 1987, Juárez & Blomquist 1993). Gas chromatographic analysis of adults of T. dimidiata, T. barberi and D. maxima showed characteristic hydrocarbon profiles, with carbon number ranging from 22 to more than 40, with a mixture of saturated straight and methyl-branched chains, with one, two, and three methyl groups, external and internally located. Methylbranched chains exhibited methyl-branching patterns consistent with previous data from other Triatominae (Juárez & Blomquist 1993, Juárez et al. 2000, 2001). However, when T. dimidiata was compared to closely related species T. pallidipennis and T. mazzotti (phyllosoma complex), differences were evident. T. dimidiata showed larger amounts of branched chains, with terminally mono, di- and trimethyl derivatives of C29, C31 and C33 accounting for 26% of the total hydrocarbon content, whereas these components were present in minor amounts in the other species (Juárez & Blomquist 1993). The major components for both species eluted quite closely at 37.9 ECL (corresponding to 3790 KI), the mass spectral identification showed large amounts of a mixture of x,x-dimethyl C37 together with x-me C38 for T. pallidipennis (Juárez & Brenner 1987) and at 37.7 ECL (ca. 3770 KI) for T. mazzotti (Juárez & Blomquist 1993); T. dimidiata showed a peak eluting at 3770 KI (x,x- dimethyl C37), although quantitatively less relevant. Sex-dependant, quantitative differences in certain hydrocarbons were found in T. dimidiata. The comparison of three selected peak ratios (R values) separated T. dimidiata males and females at the 95% level of confidence (Table III) however, both sexes were not distinguishable for T. barberi and D. maxima. Analysis at genus level based on the complete 18S rRNA gene showed the impossibility of distinguishing the genera Dipetalogaster, Panstrongylus and Triatoma (Bargues et al. 2000), however the rather simple hydrocarbon pattern of Dipetalogaster is easily differentiated from both Triatominii (Juárez et al. 2000). The analysis of triatomine wings by CGC is reliable and thus enables comparison of these data to those obtained by other techniques for specimen identification on the same insect, and collaborative work is presently addressing this subject. Among more than 130 species reported for Triatominae, the cuticular hydrocarbon structure and CGC profiles for species with major epidemiological significance, T. infestans, T. brasiliensis, Rhodnius prolixus and a number of related species, were previously reported (Juárez & Blomquist 1993, Juárez et al. 2000, 2001). A pattern of primarily quantitative rather than qualitative differences reinforces the idea that epicuticular hydrocarbons represent relatively primitive characters for these insects. T. dimidiata showed a simpler hydrocarbon fingerprint and is easily separated from species of the phyllosoma complex (Juárez & Brenner 1986, Juárez & Blomquist 1993). Within each genera examined there was an indication that species from drier regions present more complex cuticular hydrocarbon profiles than their congenerics from wetter regions. Although interspecific variations might be more relevant, the complexity of the hydrocarbon pattern of T. dimidiata was intermediate between those reported for T. tibiamaculata and T. vitticeps from high relative humidity (RH) coastal regions, and that for T. infestans from varying RH areas (Juárez et al. 2000). T. barberi of drier regions, showed a more complex pattern. D. maxima surface hydrocarbon mixture showed the largest amounts of straight chains from 60% to 67.8% of the total hydrocarbon mixture, for males and females respectively; the relative abundance of n-alkanes might be related to exposure to warm conditions.
ACKNOWLEDGEMENTS
To CJ Schofield for his critical and helpful comments, editing and review of the manuscript. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02157f3.jpg] [oc02157t4.jpg] [oc02157f2.jpg] [oc02157f1.jpg] [oc02157t2.jpg] [oc02157t1.jpg] [oc02157t5.jpg] [oc02157t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}