 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(6) September 2002, pp. 835-838 Laboratory Evaluation of Mesocyclops annulatus (Wierzejski, 1892) (Copepoda: Cyclopidea) as a Predator of Container-breeding Mosquitoes in Argentina María V Micieli+, Gerardo Marti, Juan J García
Centro de Estudios Parasitológicos
y de Vectores, Calle 2 Nº 584 (1900) La Plata, Argentina Received 1 June 2001 Code Number: oc02159
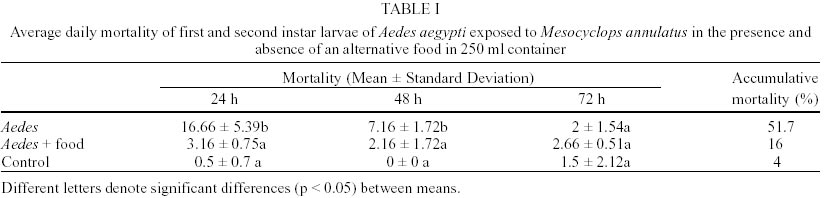
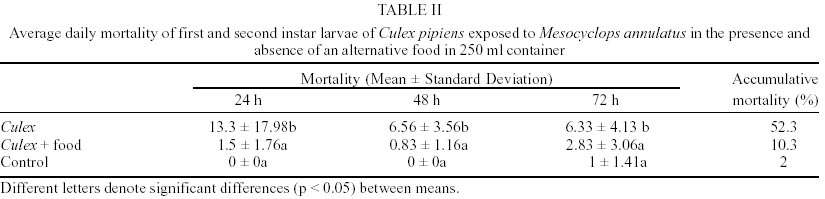
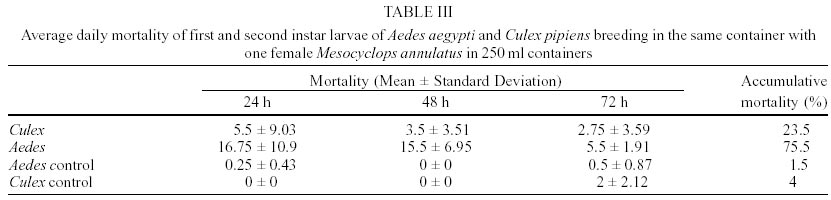
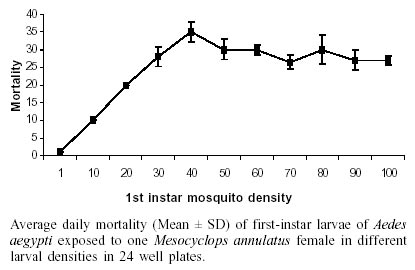
In laboratory bioassays we tested the predatory capacity of the copepod Mesocyclops annulatus on Aedes aegypti and Culex pipiens larvae. A single adult female of M. annulatus caused 51.6% and 52.3% mortality of 50 first instar larvae of Ae. aegypti and Cx. pipiens respectively, in a 72 h test period. When alternative food was added to the containers, mortality rates declined to 16% and 10.3% for Ae. aegypti and Cx. pipiens respectively. When 50 first instar larvae of each of the two mosquito species tested were placed together with a single adult female of M. annulatus, mortality rates were 75.5% for Ae. aegypti larvae and 23.5% for Cx. pipiens larvae in a three day test period. Different density of adult females of M. annulatus ranged from 5 to 25 females produced mortality rates of Ae. aegypti first instar larvae from 50% to 100% respectively. When a single adult female of M. annulatus was exposed to an increasing number of first-instar Ae. aegypti larvae ranging from 10 to 100, 100% mortality was recorded from 1 to 25 larvae, then mortality declined to 30% with 100 larvae. The average larvae killed per 24 h period by a single copepod were 29.
Key words: copepod - Mesocyclops annulatus - Aedes aegypti - Culex pipiens - Argentina Aedes aegypti L., eradicated from Argentina in 1963, has reinfested this country as far as south of Buenos Aires city and dengue is certainly emerging as an important disease in Argentina (Avilés et al. 1999). Vector control with insecticides may cause environmental damage and mosquito resistance (Breakley et al. 1984). Source reduction is appropriate in controlling Ae. aegypti larvae but some breeding sites are not amenable to this method. The use of biological control against Ae. aegypti is an alternative control measure that should be considered. Culex pipiens L. is also one of the major mosquito pest in Argentina and with Ae. aegypti are the two most common container-breeding mosquitoes. Among all the natural enemies mentioned in the literature as potential candidates for mosquito control (Jenkins 1964, Roberts & Strand 1977, Roberts & Castillo 1980, Roberts et al. 1983), predacious copepods are promising candidates for biological control of container-breeding mosquitoes (Marten 1990). They are particularly effective predators of mosquito larvae. In addition, copepods do not depend on the supply of mosquito larvae for food. Copepods can feed on algae, protozoa, rotifers and others arthropod larvae. Various species of Mesocyclops, Macrocyclops, Megacyclops and Acanthocyclops have been tested in a variety of Aedes-breeding habitats (Riviere & Thirel 1981, Suarez et al. 1984, Riviere et al. 1987, Kay et al. 1992, Marten et al. 1994) with promising results. Laboratory evaluation of these potential agents of biological control is an essential preliminary step toward selection of an appropriate candidate for field releases. In this study we reported the predatory behavior of a common temporary pool copepod, M. annulatus (Wierzejski, 1892). Laboratory trials were conducted to evaluate the effectiveness of this predaceous cyclopoid for control of Ae. aegypti and Cx. pipiens larvae in domestic containers in La Plata, Argentina. MATERIALS AND METHODS
Collection and culture of M. annulatus - Copepods were collected using a net (100 µm mesh) through the margins of transient freshwater ponds near La Plata city, Buenos Aires province, Argentina (34º 51' 07" S, 58º 57' 30" W). Copepods were identified according to Reid (1985) and reconfirmed by Dr S Menú Marquez, University of Buenos Aires, Argentina. Individual gravid females of M. annulatus were placed in containers containing 200 ml of dechlorinated tap water, and added protozoan and algae as food. Copepod larvae were placed in containers with 2000 ml of dechlorinated water and incubated at 26 ± 2°C to adult stage. Source of mosquito larvae - Larvae of Ae. aegypti and Cx. pipiens used in this study were obtained from colonies maintained at Centro de Estudios Parasitológicos y de Vectores, La Plata, Argentina. Larvae were kept in plastic pans (40 x 30 x 6 cm) with dechlorinated tap water until pupation and they were fed on powdered chicken food. Pupae were placed in screened cages for adult emergence. Adults of both species were fed on 10% sucrose solution and a restrained chicken was offered as a blood source twice a week. Cx. pipiens females were allowed to lay eggs in a container half filled with water. A strip of filter paper was added inside the perimeter of the container for oviposition of Ae. aegypti females. Predation trials - Experiment 1: predatory capacity of M. annulatus on first-instar larvae of Ae. aegypti and Cx. pipiens was determined. A single M. annulatus adult female was placed with 50 first-instar mosquito larvae (24 h old) in a 250 ml plastic container (8 x 8 x 5 cm) containing 100 ml of dechlorinated tap water at 26ºC. This experiment was conducted over 3 days, and number of surviving larvae on each day was recorded. To assess copepod feeding in the presence of an alternate food source, identical trials were conducted under similar conditions as described above with addition of 10 ml algal-protozoan suspension per container from an eutrophic pond. Experiment 2: this experiment was carried out for comparison of the effectiveness of M. annulatus as predators of Ae. aegypti and Cx. pipiens larvae living in the same container. Fifty larvae of each mosquito species and one adult female copepod were introduced in containers containing 100 ml of dechlorinated tap water. Experiment 3: it was designed to determine the number of adult copepods needed to consume a large number of first instar Ae. aegypti larvae. Five, 10, 15, 20 and 25 M. annulatus adult female were introduced in containers (11 x 11 x 12 cm) with 100 first instar larvae of Ae. aegypti and 700 ml of dechlorinated water. Surviving larvae were counted at 24 h. Experiment 4: predation capacity of individual M. annulatus female was determined in relation to different Ae. aegypti larval densities. Densities of first instar larvae of Ae. aegypti from 1 to 29 (increasing by 1) and from 30 to 100 (increasing by 10) were placed in 24 well plates (1.6 cm diam.) with 1 M. annulatus adult female and 3 ml of dechlorinated tap water per well. Larval mortality was determined at 24 h. Similar tests were conducted in 250 ml containers with 100 ml of water. These experiments were conducted at 26 ± 1 °C under a photoperiod of 12:12 (L:D) h. Six replicates and two controls (containers with larvae without copepods) were conducted for each of the four experiments. Statistical analysis - Mean number of killed larvae was compared between treatment and time by a two-way ANOVA. Multiple comparisons were tested by Tukey HSD test (Sokal & Rohlf 1995). In order to homogenize the variances, killed larvae numbers (n) were log (n + 1) transformed before to the analysis. RESULTS
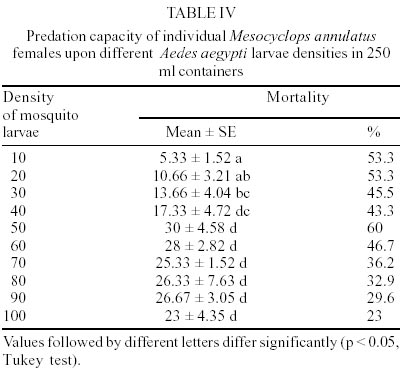
M. annulatus preyed on first and second instar larvae of both mosquito species, Ae. aegypti and Cx. pipiens. However, predation on Ae. aegypti larvae was affected significantly by presence of alternative food (F = 34.03, df = 3,36, P < 0.001). In the absence of alternative food, significant larval mortality was observed within 24 h and 48 h. Larval mortality was not significantly different between treatment and control at 72 h (Table I). By day 3, an average of 51.7% larval reduction was recorded in containers with copepods. In presence of alternative food, no significant larval mortality relative to controls was detected (Table I). Total mortality of Ae. aegypti larvae in containers with alternative food averaged 16%. Predatory effectiveness of M. annulatus against first and second instar Cx. pipiens was affected significantly by presence of alternative food (F = 44.8, df = 3,36, P < 0.001). In absence of alternative food, daily larval mortality was significantly different from controls at 24, 48 and 72 h (Table II). Mortality was 52.3% during the 3 day test period. In presence of an alternate food source, no significant larval mortality relative to controls was detected. Total mortality was 10.3% for the 72 h test period. When 50 first instar larvae of Ae. aegypti and 50 first instar larvae of Cx. pipiens were added in containers with 1 copepod, 53.5% of Ae aegypti and 11% of Cx. pipiens larvae were killed at 24 h. Total larvae mortality was 23.5% in Cx. pipiens and 75.5% in Ae. aegypti larvae for the 72 h test period. This difference was significant (F = 23.55, df = 3,36, P < 0.001). Densities of 5, 10, 20 and 25 adult females of M. annulatus placed in 700 ml containers with 100 first-instar larvae of Ae. aegypti produced 49.7%, 84%, 97% and 100% mortality respectively during 72 h test period. When a single M. annulatus female was placed with larval densities of 1 to 29, 30, 40, 50, 60, 70, 80, 90 and 100 in 24 well plates with 3 ml per well, 100% mortality was obtained from 1 to 25 larvae. The average number of larvae killed by a single copepod female was 29 (26-35) (Figure). When a similar experiment was conducted in containers with 100 ml, at densities from 40 to 100 larvae per container no significant differences were observed on mortality rates. Mortality rates on densities of 10, 20 and 30 larvae per container were significantly different with densities larger than 30, 40 and 50 respectively (Table IV). DISCUSSION
In this study, we demonstrated that adult female of M. annulatus preyed on larvae of Ae. aegypti and Cx. pipiens younger than 72 h (early second instar). In addition, presence of alternative food in containers with mosquito larvae and M. annulatus produced from 3 to 5 fold reduction on predation effectiveness. Our results suggest that M. annulatus did not have a feeding preference for mosquito larvae in containers if algae and protozoan were available. Reduction of predation rates of copepods on mosquito larvae in the presence of abundantly alternative food has been reported by previous authors (Rivière et al. 1987, Laird 1988, Andreadis & Gere 1992). Some species of Mesocyclops have been previously reported as non-efficient predators of Culex because of the larvae ability to escape capture by copepods (Rivière et al. 1987, Marten et al. 1989, Marten 1990). This study also reports reduced mortality on Culex compared to Aedes larvae. In experiments carried out in 700-ml artificial containers, which simulated common breeding sites for Ae. aegypti and Cx. pipiens such as flower vases in cemeteries, we found that the addition of an average of 25 M. annulatus adult female produced 100% control on first instar Ae. aegypti larvae. There was no significant differences in mortality when Ae. aegypti and Cx. pipiens larvae were tested separately, in contrast when both culicid species were simultaneously offered to copepod a selective consumption of Ae. aegypti larvae was registered. Larvae of Ae. aegypti are benthic feeder while Cx. pipiens larvae are surface feeder (Laird 1988). Although most of the Mesocyclops in lakes are known as planktonic organisms (Laird 1988), our observations on feeding behavior of M. annulatus indicated that this copepod species remained at the bottom of the containers most of the time. Feeding behavior of these mosquito and copepod species may lend to increase encounter opportunities with Ae. aegypti larvae by M. annulatus resulting in the differential predation observed. Previous studies have shown the importance of morphological and behavioral characteristics other than prey size as defense mechanisms from predation by cyclopoids (Kerfoot 1978, Williamson 1983). Prey density and container size were also important components in the predation effectiveness of M. annulatus. When a single copepod was placed in a small container, it killed on average of 29 larvae in 24 h, independent of the larvae density. When larger containers were used, similar numbers of larvae were killed only at densities larger than 50 larvae. We speculate that M. annulatus was not able to reach the daily consumption of 29 larvae averaged at densities lower than 50 larvae because of its lack of ability to locate the prey in larger containers. These laboratory studies indicate that although M. annulatus seems promising as a biocontrol agent, similar trials should be done with other local species prior to choice of a candidate for field testing.
ACKNOWLEDGMENTS
To Dr Silvina Menú Marquez (Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina) for reconfirmation of copepods identity. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02159t4.jpg] [oc02159f1.jpg] [oc02159t3.jpg] [oc02159t2.jpg] [oc02159t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}