 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(6) September 2002, pp. 857-864 Changes in Nuclear Phenotype Frequencies Following Sequential Cold Shocks in Triatoma infestans (Hemiptera, Reduviidae) Silvana GP Campos, Vera Lúcia CC Rodrigues*, Maria Luiza S Mello/+
Departamento de Biologia Celular,
Instituto de Biologia, Universidade Estadual de Campinas, 13084-971 Campinas,
SP, Brasil *Sucen, Mogi-Guaçu, SP, Brasil This paper received financial support from Fapesp (grant no. 99/2547-8), Capes and CNPq. This study was part of a thesis presented by SGPC to the Instituto de Biologia, Unicamp, in partial fullfilment of the requirements for a Master degree. Received 7 December 2001 Code Number: oc02164
The nuclear phenotypes of Malpighian tubule cells in fifth instar nymphs of Triatoma infestans, one of the most important vectors of Chagas disease, were studied following sequential shocks at 0ºC, separated by intervals of 8 h and 24 h at 30ºC, under conditions of moderate fasting and full nourishment. The insects pertained to colonies reared in the laboratory and originated from domestic specimens collected in the Brazilian states of São Paulo (north) and Minas Gerais (south). Since nuclear phenotypes in this species are affected by single cold shocks, it was expected that these phenotypes could also be changed by sequential shocks. Nuclear phenotypes indicative of mechanisms of cell survival (nuclear fusion and heterochromatin decondensation) and cell death (apoptosis and necrosis) were observed concomitantly in all the conditions tested. Nuclear fusion and heterochromatin decondensation were not found relevant for the presumed acquisition of the cold-hardening response in T. infestans. The decreased frequency of apoptosis and necrosis following sequential cold shocks including under fasting conditions, indicated that tolerance to sequential cold shocks occurred in T. infestans of the mentioned origin.
Key words: cold shocks - sequential shocks - nuclear phenotypes - Triatoma infestans
Triatoma infestans, a blood-sucking insect, is one of the most important vectors of Chagas disease. Survival, molting rate and chromatin supraorganization in this species are affected by stressing agents such as fasting, gamma-radiation, heavy metals and heat and cold shocks (Álvares-Garcia 1988, Mello 1989, Rodrigues et al. 1991, Dantas & Mello 1992, Mello et al. 1995, 2001, Campos et al. 2002). T. infestans is tolerant to a single shock at 0°C for up to 12 h, or to sequential cold shocks, regardless of the nutritional state of the specimens (Campos et al. 2002). The molting rate of T. infestans nymphs may be affected by cold shocks, the extent of which depends on the nutritional state of the specimens. Fasting enhances tolerance to sequential cold shocks, compared to the fully nourished state, provided the interval at 30°C between sequential shocks at 0°C is no longer than 8 h (Campos et al. 2002). The cold-hardening response in T. infestans may have favoured its survival in areas of South America which experience low temperatures, although this insect relocates predominantly in association with human habitats (Campos et al. 2002). Single cold shocks at 0°C for 1 h or 12 h induce changes in the nuclear phenotypes of the Malpighian tubules of T. infestans (Dantas & Mello 1992). The nuclei of this species normally show a single or several conspicuous chromocenters represented by heterochromatic A, B, and C autosomes plus sexual chromosomes. However, phenotypes indicative of cell survival (unravelled heterochromatin, nuclear and cell fusion) and cell death (apoptosis, necrosis) strategies are observed concomitantly especially in response to heat and cold shock stress (Dantas & Mello 1992, Mello et al. 2001). Since the nuclear phenotypes of T. infestans Malpighian tubules are affected by single cold shocks, changes in the nuclear phenotype frequencies would also be expected following sequential cold shocks. In this study, we examined the effect of sequential cold shocks on the nuclear phenotypes of Malpighian tubules in fasted and fully-nourished T. infestans. MATERIALS AND METHODS
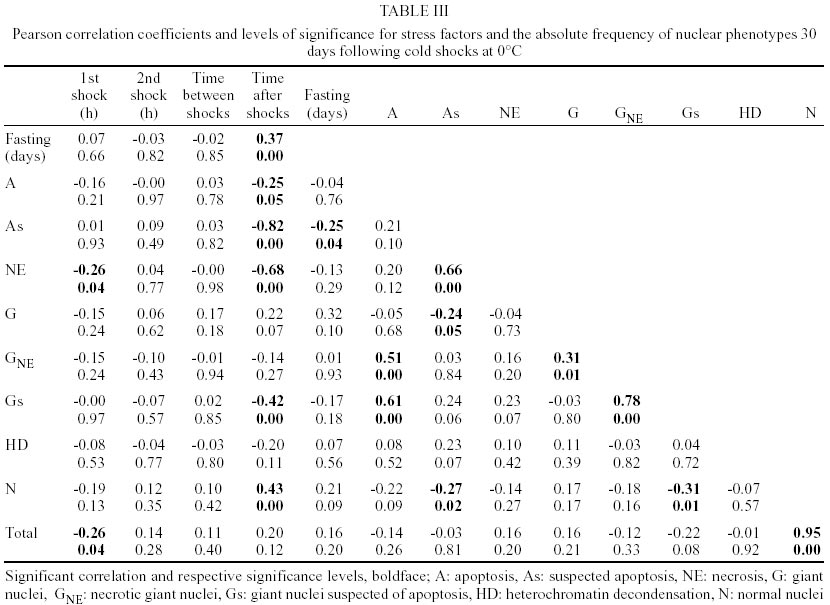
Fifth instar nymphs of T. infestans Klug (Hemiptera, Reduviidae) reared at 30°C and 80% relative humidity in the laboratory at Sucen (Mogi-Guaçu, State of São Paulo) were used. The specimens pertained to colonies reared in Sucen since 1975 and originated from domestic insects collected in the Brazilian states of São Paulo (north) and Minas Gerais (south) and, since then, periodically crossed with specimens collected in the south of Minas Gerais. The insects were fed once a week on hen blood or were fasted from 15 days before to 30 days after the shocks. Immediately after a shock at 0ºC for 1 h, the insects were returned to 30°C; 8 h and 24 h later, they were subjected to an additional shock at 0°C which lasted 12 h. A temperature of 0°C was chosen based on previous results (Rodrigues et al. 1991, Campos et al. 2002). Nymphs subjected to a single shock at 0°C for 1 h or 12 h were controls 1 and 2, respectively. A description of data for nymphs maintained at 30°C, a temperature used to rear T. infestans in the laboratory since 1980 (Rodrigues et al. 1991), has been provided. Following shocks, the nymphs were returned to 30°C. Malpighian tubule preparations were obtained 2 and 30 days after single or sequential cold shocks. The organs from at least three specimens were used for each tested condition. Whole Malpighian tubules were mounted on glass slides, fixed in ethanol-acetic acid (3:1, v/v) for 1 min, rinsed in 70% ethanol for 5 min, and air dried at room temperature. The material was then subjected to the Feulgen reaction, with hydrolysis in 4 M HCl at 25°C for 65 min. The Feulgen-stained material was rinsed in sulfurous and distilled water, air dried, cleared in xylene, and mounted in Canada balsam. The total number of Feulgen-stained Malpighian tubule epithelial cell nuclei and the number of different nuclear phenotypes were counted in each specimen. Photomicrographs were obtained using a Zeiss Axiophot 2 microscope (Oberkochen, Germany). Pearson product moment correlation coefficients were used to assess the degree of linear relationship between multiple columns of pairs of variables (correlation matrix) for fully-nourished and moderately fasted specimens (Minitab12TM statistical software, State College, USA). The Pearson correlation coefficient is the most widely used measure of association between two variables, reflecting the strength of association between them (Forthofer & Lee 1995). This statistical test has been largely used in studies on linear correlation between heat or cold shocks and the relative frequency of nuclear phenotypes in Panstrongylus megistus (Garcia et al. 2000a, b), and between heat shock, nutritional conditions and the relative frequency of nuclear phenotypes in T. infestans (Mello et al. 2001). RESULTS
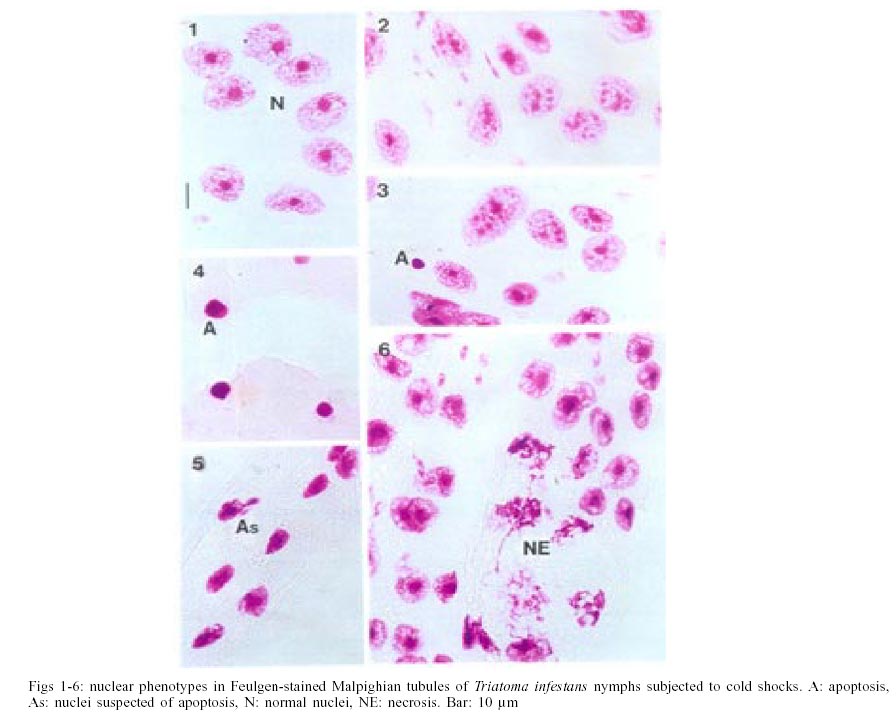
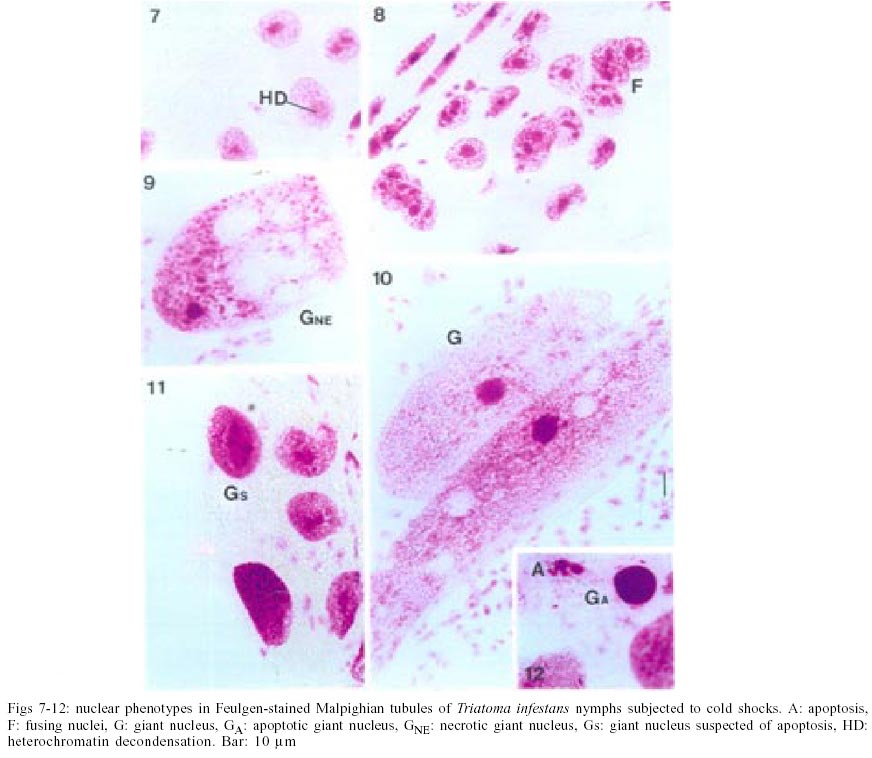
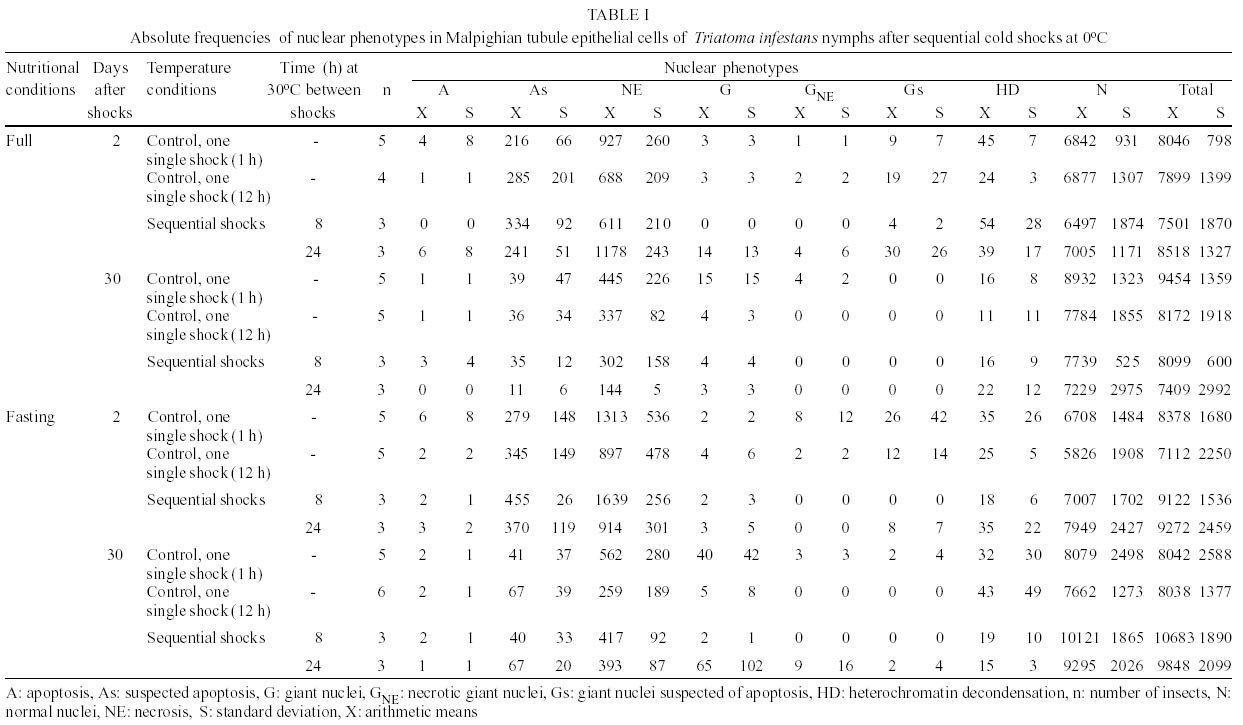
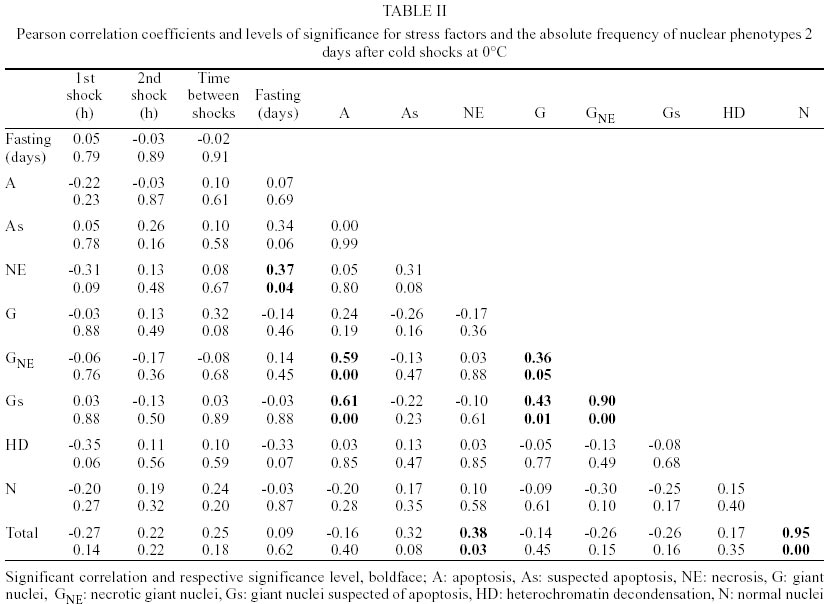
The most common nuclear phenotypes in Malpighian tubules of T. infestans contained one or several heterochromatic bodies (chromocenters) amidst euchromatin (Figs 1, 2), regardless of the type of cold shock, nutritional state or time following the shock. In altered nuclear phenotypes, apoptotic nuclei (A) (Figs 3, 4), nuclei suspected of apoptosis (As) (Fig. 5), necrotic nuclei (NE) (Fig. 6), nuclei with heterochromatin decondensation (HD) (Fig. 7) and giant nuclei (G) (Figs 8, 10) were observed. In some cases, the G showed signs of necrosis (GNE) or As (GA and Gs) (Figs 9-12). The A contained evenly distributed condensed chromatin; A vesicles were sometimes present. As nuclei were those in which the euchromatin stained as deeply as heterochromatin with the Feulgen reaction, showing a chromatin condensation pattern similar to that for other cell systems in which a decrease in Feulgen-DNA amounts precedes the drastic DNA losses typical of advanced stages of A (Maria et al. 2000). The necrotic nuclei showed various stages of nuclear disruption. HD affected the chromocenters either partially or totally. The G were generated by nuclear and/or cell fusion (Fig. 8) (Mello 1989). Table I shows arithmetic means (X) and standard deviations (S) for the frequency of the various nuclear phenotypes and the total number of nuclei of the Malpighian tubule cells in fully-nourished and fasted specimens. Since individual variations in the number of nuclei are common in the Malpighian tubules of reduviids (Andrade & Mello 1987, Dantas & Mello 1992, Mello et al. 1995), the arithmetic mean may not be representative of the respective group. That occurred in cases where a certain phenotype was observed in a few nuclei (A, G, GNE, Gs). Pearson correlation coefficients were calculated for each of the following pairs of variables: frequency of nuclear phenotypes A vs As, A vs NE, A vs G, A vs GNE, A vs Gs, A vs HD, A vs N, As vs NE, As vs G, As vs GNE, As vs Gs, As vs HD, As vs N, NE vs G, NE vs GNE, NE vs Gs, NE vs HD, NE vs N, G vs GNE, G vs Gs, G vs HD, G vs N, GNE vs Gs, GNE vs HD, GNE vs N, Gs vs HD, Gs vs N, HD vs N; frequency of each nuclear phenotype vs total number of nuclei; frequency of each nuclear phenotype and of total nuclei vs number of days following fasting; frequency of each nuclear phenotype and of total nuclei vs time (hours) between shocks; frequency of each nuclear phenotype and of total nuclei vs time (days) after shocks (Tables II, III). If one variable tended to increase as the other decreased, the correlation coefficient was negative. Conversely, if the two variables tended to increase (or decrease) together, the correlation coefficient was positive. The p-value for each correlation coefficient was supplied by the statistical software. Correlation values significant at P < 0.05 appear highlighted in Tables II and III. In all cases, normal nuclear phenotypes were the most common, followed by NE. The latter was significantly more frequent under moderate fasting in specimens examined two days after shocks (positive correlation, P0.04) (Table II), but decreased thereafter (non significant correlation) (Table III). The total number of nuclei did not decrease significantly with fasting (Tables II, III). A was rarely observed in the various groups analyzed, especially 30 days after the shocks (Table I). In only a few specimens did the A represent > 0.20% of the total cell nuclei. G and nuclei with HD were also rare in the various groups. Typical A involving G (Fig. 12) were observed in a few cells of just one specimen and not considered in the statistical approach. The frequency of nuclei with HD did not vary significantly in the various experiments (Tables II, III). The frequency of A and As decreased significantly with increasing time after the shocks (negative correlation significant at P0.05 and P0.00, respectively) (Table III). A in normal-sized nuclei (A) correlated significantly with the frequency of G showing necrosis (GNE) or As (Gs) following cold shocks (P0.00, Tables II, III). There was no correlation between total number of nuclei or a specific nuclear phenotype and the sequential cold shock conditions, or the time at 30°C between shocks at 2 and 30 days after the shocks (Tables II, III). There was a significant decrease in the number of nuclei with NE and in the total number of nuclei after single shocks (negative correlation significant at P0.04) (Table III). Only NE correlated positively (P0.04) with the fasting state and with the total number of nuclei (P0.03), 2 days after the cold shocks (Table II). However, the frequency of NE decreased significantly with increasing time after the shocks (negative correlation significant at P0.00) (Table III). DISCUSSION
Normal nuclear phenotypes, as well as nuclear phenotypes indicative of mechanisms of cell survival (nuclear fusion and HD) (Wigglesworth 1967, Mello 1989, Mello et al. 2001) and cell death (A and NE) occurred concomitantly in the Malpighian tubules of T. infestans under all tested situations. These phenotypes have been reported after single cold and heat shocks (Dantas & Mello 1992, Mello et al. 2001), as well as with other stress conditions such as gamma radiation, long fasting periods, and heavy metals (Álvares-Garcia 1988, Mello 1989, Mello et al. 1995). In heat shock tests, HD is more relevant for cell survival in fully-nourished nymphs, whereas nuclear and cell fusions are more important in fasted specimens (Mello et al. 2001). In the present study, neither nuclear and/nor cell fusions or HD were important for acquisition of the cold-hardening response. Nuclear fusion, which generates G in several reduviid species, especially after long periods of fasting (Wigglesworth 1967, Mello & Raymundo 1980, Andrade & Mello 1987, Mello 1989), increased 30 days after sequential cold shocks in fasted T. infestans. Even so, the presence of G was occasional and not sufficient to significantly affect the total number of nuclei estimated under fasting plus cold shock conditions. When cell/nuclear fusion was induced as part of a cold-hardening response in T. infestans, many G showed cell death morphologies, indicating that this approach was not particularly efficient. HD was not a significant surviving strategy following cold shocks. Indeed, HD is not required for cell survival following cold shocks in T. infestans, in contrast to Panstrongylus megistus, a reduviid species which is more sensitive to cold shock and acquires cold-shock tolerance following sequential cold shocks (Garcia et al. 1999, 2000a, 2001). A and NE are forms of cell death with typical morphological aspects in the Malpighian tubules of reduviids (Dantas & Mello 1992, Mello et al. 1995, 2001, Garcia et al. 2000a, b). In our study, the incidence of A was low, especially following sequential cold shocks, and was independent of the number and duration of shocks, of the interval at 30ºC between shocks, and of the nutritional state. The frequency of A decreased in the 30-day period post-shock. This observation was presumably related to the cold-tolerance response developed by T. infestans. NE, a form of cell death resulting from severe cell damage, was observed in all specimens analyzed. Cold shocks apparently injure the Malpighian tubule cells of T. infestans, the effect being greater in specimens with a higher nuclear frequency. However, NE increased only in fasted specimens two days after the sequential shocks. The decrease in the frequency of cell death forms following sequential cold shocks including under fasting conditions, indicated the acquisition of cold-shock tolerance in T. infestans. Indeed, in terms of insect survival, T. infestans is tolerant to single or sequential shocks at 0ºC, regardless of the nutritional state of the specimens, whereas in terms of molting rate, fasting elicits a protective action, in addition to that induced by cold shocks (Campos et al. 2002). Since the specimens of T. infestans analyzed here originated from insects collected in the State of São Paulo and south of the Minas Gerais, their source is concerned with the hypothesis of the species coming from Argentina (Aragão 1971, Silva 1986, Monteiro et al. 1999). According to Noireau and his co-workers (2000), domestic populations of T. infestans from Brazil and Argentina differ from the sylvatic populations of this species occurring in the Andean valleys of Bolivia. Even so, these evolutionary units may still share a common background (Dujardin et al. 1999) and as such a tolerance to cold shocks that may have facilitated the survival of this species at low temperatures in association with high altitudes in the Andes mountains (Usinger et al. 1966) would also be a characteristic of the T. infestans populations found in Brazil. Involvement of heat-shock proteins and other mechanisms including genetic variation in metabolic traits (Clark & Fucito 1998), may be responsible for the cold-hardening response in T. infestans. In contrast to the decrease in the total number of nuclei in the Malpighian tubules of T. infestans after very long periods of fasting (Mello 1989), the lack of change observed in the present study may reflect the relatively mild fasting protocol used. REFERENCES
The following images related to this document are available:Photo images[oc02164t2.jpg] [oc02164t1.jpg] [oc02164f1-6.jpg] [oc02164t3.jpg] [oc02164f7-12.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}