 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(6) September 2002, pp. 901-903 SHORT COMMUNICATION
Time Course of in Vitro Maturation of Intra-erythrocytic Malaria Parasite: a Comparison between Plasmodium falciparum and Plasmodium knowlesi SD Srinivas+, SK Puri
Division of Parasitology, Central
Drug Research Institute, Lucknow, India Financial support from Senior Research Fellowship, Council of Scientific & Industrial Research, New Delhi, India. Received 28 August 2001 Code Number: oc02173
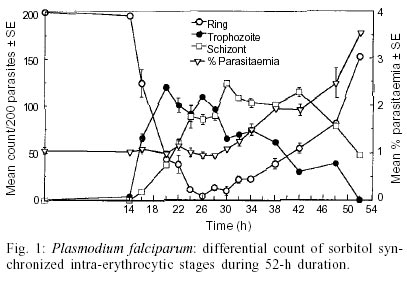
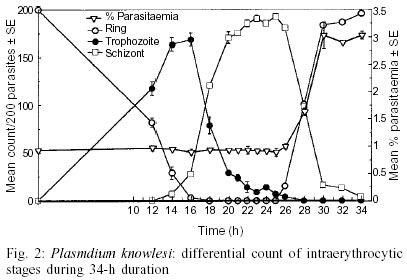
The schizont maturation assay for in vitro drug sensitivity tests has been a standard method employed in the global baseline assessment and monitoring of drug response in Plasmodium falciparum. This test is limited in its application to synchronous plasmodial infections because it evaluates the effect of drug on the maturation of parasite especially from ring to schizont stage and therefore synchronized P. falciparum cultures are required. On the other hand, P. knowlesi, a simian malaria parasite has a unique 24-h periodicity and maintains high natural synchronicity in monkeys. The present report presents the results of a comparative study on the course of in vitro maturation of sorbitol synchronized P. falciparum and naturally synchronous P. knowlesi. Ring stage parasites were incubated in RPMI medium supplemented with 10-15% pooled homologous serum in flat-bottomed 96-well micro plates using a candle jar at 37°C. The results suggest that the ideal time for harvesting the micro-assay plates for in vitro drug sensitivity test for sorbitol-synchronized P. falciparum and naturally synchronous P. knowlesi are from 26 to 30 h and from 22 to 25 h, respectively. The advantages of using P. knowlesi in chemotherapeutic studies are discussed.
Key words: Plasmodium falciparum - Plasmodium knowlesi - schizont maturation test - in vitro drug sensitivity test - sorbitol synchronization The efficacy of an antimalarial drug depends on its ability to inhibit or interrupt essential life functions of the malaria parasite. The resurgence of malaria in various areas of the world, combined with the expansion of chloroquine resistant infections in the late 60s, focused attention on the development of in vitro tests which would permit the objective measurement of the sensitivity of human plasmodia to antimalarial drugs. After the achievement of continuous cultivation of Plasmodium falciparum in 1976 in appropriate media (Trager & Jensen 1976, Haynes et al. 1976), Rieckmann et al. (1978) adapted this methodology to produce a microculture procedure for the assessment of schizont maturation in a short-term test using chloroquine and mefloquine. Schizont maturation tests are essentially limited in their application to synchronous plasmodial infections or to those in which only one asexual growth form is generally found in the peripheral blood as the test studies the effect of drug on the maturation of parasite from ring to schizont stage. In human malarias this fact restricts their use to P. falciparum infection as only ring stage parasites are found in peripheral blood due to sequestration of erythrocytes containing advanced stages in capillaries. In this case, the schizont maturation test is widely applied and the microculture procedure has been the standard method employed in the global baseline assessment and monitoring of drug response (Lopez-Antunano & Wernsdorfer 1979, WHO 1984, Draper et al. 1985, Afari et al. 1993, Mberu et al. 2000). Since schizont maturation tests require uniform ring stage parasites, the asynchronous P. falciparum cultures are synchronized by chemical treatment. In this report we made an attempt to study the time course of in vitro maturation of sorbitol synchronized ring stage parasites of a culture adapted P. falciparum strain and compare these results with those obtained with in vitro maturation of a monkey malaria parasite, P. knowlesi, which has a unique quotidian periodicity and high natural synchronicity infection in monkeys (Collins 1988, Cogswell 2000). These features of the simian parasite add to the advantages of this species for wider application in chemotherapeutic studies. A culture adapted strain (FCD-3) of P. falciparum obtained from the International Center for Genetic Engineering and Biotechnology, New Delhi, India and W1 strain of the simian parasite, P. knowlesi which was originally provided by Prof. PC Garnham (England) in 1976 and has been maintained at Central Drug Research Institute via blood induced passages in rhesus monkeys as well as cryopreservation in liquid nitrogen were used in the present study. Blood induced passages were carried out by intravenous inoculation of healthy rhesus monkeys with 1 ml of infected rhesus blood in citrate anticoagulant solution (pH 7.4) from another infected monkey or 1 ml of revived parasites that were cryopreserverd (Rowe et al. 1968) in liquid nitrogen. P. falciparum was maintained continuously using a simple candle jar method (Trager & Jensen 1976) in RPMI1640 medium containing 25 mM HEPES (Sigma, USA) and supplemented with 0.23% sodium bicarbonate, 1% D-glucose and 10-15% heat inactivated O+ human serum. Synchronization of the parasites to uniform ring stages was achieved using 5% aqueous D-sorbitol (Lambros & Vanderberg 1979). Briefly, culture contents were centrifuged to pellet the cells. The supernatant media was discarded and packed cells were suspended in 5 times volume of 5% aqueous D-sorbitol and allowed to stand for 15 min. The cells were washed twice with RPMI medium without serum to remove sorbitol and appropriate volume of red blood cells (RBCs) were added to obtained the required parasitaemia level. Highly synchronous P. knowlesi ring stage parasites were obtained from an experimentally infected rhesus monkey. Parasitized blood was drawn by venepuncture in citrate solution and was washed free of plasma and white blood cells with RPMI medium. For in vitro schizont maturation assay with either of the parasites, 10-15% heat inactivated pooled homologous serum was added to RPMI medium. The time course of in vitro schizont maturation of parasites was monitored in flat bottomed 96 well culture plates (Tarsons). Parasitized ring stage erythrocytes were diluted with fresh uninfected homologous erythrocyes to obtain parasitaemia between 0.5 to 1%. They were diluted using serum containing RPMI 1640 medium to 6% haematocrit. A total of 200 µl aliquots of the culture were added to each well. The plates were covered and incubated in a candle jar at 37°C. At different sequential intervals, the plates were removed and thin smears from the settled RBCs were made from triplicate wells. The smears were stained with Giemsa stain and the differential stages (ring, trophozoites, schizonts) were microscopically recorded for 200 asexual parasites in each smear. Fig. 1 represents the differential counts of P. falciparum parasites from samples taken at different times during a 52-h period, initiated with sorbitol synchronized intra-erythrocytic ring stages. The proportion of early:late rings obtained after sorbitol synchronization was 148:52 out of 200 parasites. Preschizonts (representing schizonts with 2-3 nuclei) and schizonts began to appear in culture from the 16th h onwards gradually increasing to a maximum of 34:90 (total = 124) ratio at 30 h. At 32 h, reinvasion of uninfected red cells started, giving an average count of 22 early rings per 200 parasites, which gradually increased at the expense of schizonts to a maximum of 152/200 parasites at 52 h. Consequently, the parasitaemia started increasing from 32 h onwards reaching a maximum of 3.5% at 52 h. Sorbitol treatment is employed to synchronize P. falciparum cultures. While the uninfected erythrocytes are impermeable to sorbitol, the changes caused in the erythrocyte permeability by the developing parasites allow sorbitol to enter erythrocytes causing lysis of cells. It should be noted that sorbitol exposure has not been shown to have any effect on the intracellular parasite growth (Lambros & Vandererg 1979, Ginsburg et al. 1987, Nakazawa et al. 1995). This treatment results in cell suspensions having only ring stage parasites estimated to be up to 18 h old (Jensen 1988). Therefore, the maximum number of schizont formation would vary considerably between 22 and 30 h depending on the number of late rings present in culture soon after synchronization of parasites by sorbitol treatment. Thus, the ideal time for harvesting microassay plates for sorbitol synchronized P. falciparum cultures could be between 26 and 30 h. Similarly, Rieckmann et al. (1978) found 24-28 h to be the ideal time to harvest the test when P. falciparum parasites obtained directly from patients were used. However, there is evidence that the incubation time for in vitro microtest varies with the parasite strain/isolate of chloroquine resistant P. falciparum and the addition of different serum types in culture (Smrkovski et al. 1983, Inaba et al. 2001). Differential counts of P. knowlesi parasites, from samples taken at different times during a 34-h period, initiated with naturally synchronous intra-erythrocytic ring stages are shown in Fig. 2. The results show the formation of a few two nucleated schizonts as early as 14 h. Multinucleated schizonts appeared at 16 h. There was a gradual increase in the number of schizonts reaching a maximum of 189 per 200 parasites by the 25th h. Re-invasion of uninfected erythrocytes started from the 26th h as indicated by the appearance of ring stage parasites, which gradually increased to 196/200 parasites at 34 h. This occurred due to the rupture of schizonts and consequently the mean schizont numbers gradually declined to 4 per 200 parasites at 34 h. Apparently, the parasitaemia level showed no major change up to 25 h but gradually started increasing to a maximum of 3.03% at 34 h as seen in Fig. 2. Thus, the ideal time for harvesting the plates for drug sensitivity assay for P. knowlesi would be between 22 and 25 h. Initiation of in vitro P. knowlesi cultures with early ring stage parasites resulted in maturation of maximum number of schizonts in vitro at the 25th h. This indicates a slight slower rate of intracellular growth as compared to the in vivo situation where the asexual schizogony is known to be complete in exactly 24 h. The monkey malaria parasite shows a sharp peak in numbers of the schizont as compared to the sorbitol synchronized P. falciparum cultures. Moreover, we found that the continuous cultivation of this simian parasite did not have considerable effect on the synchronicity of the parasite in culture. In recent years, the ex-vivo and in vitro studies to understand the pharmacokinetic-pharmacodynamic relationship of antimalarial drugs using both animal and human models have gained importance (Rieckmann et al. 1996, Sowunmi & Oduola 1998, Ubalee et al. 1999). Such studies not only increase the value of in vitro culture systems of malaria parasites in chemotherapeutic applications, but the availability of a P. knowlesi in vitro model would provide an opportunity to compare in vitro, in vivo and ex vivo response of potential antimalarial drugs against the same parasite species. Although rhesus monkeys models are widely used in malaria research, the use of these monkeys is limited in the `new world' because of the high importation and maintenance cost. However, collaborative research efforts with the `old world' sustain the continued use of rhesus monkeys. ACKNOWLEDGEMENTS
To the Director of Central Drug Research Institute for providing facilities. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02173f2.jpg] [oc02173f1.jpg] |
| |||||||||

{kind=link}
{kind=link}