 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(Suppl. I) October 2002, pp. 11-14 Experimental Evidence and Ecological Perspectives for the Adaptation of Schistosoma mansoni Sambon, 1907 (Digenea: Schistosomatidae) to a Wild Host, the Water-rat, Nectomys squamipes Brants, 1827 (Rodentia: Sigmodontinae) Paulo Sérgio D'Andrea/+, Fabiano Araujo Fernandes, Rui Cerqueira*, Luis Rey
Laboratório de Biologia e
Controle da Esquistossomose, Departamento de Medicina Tropical, Instituto Oswaldo
Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil *Laboratório
de Vertebrados, Departamento de Ecologia, Universidade Federal do Rio de Janeiro,
Rio de Janeiro, RJ, Brasil This work was supported by Fiocruz, Faperj, FUJB, Ministério do Meio Ambiente/Probio and CNPq. Received 18 June 2002 Code Number: oc02178
Due to the semi aquatic habits and the overlap of the geographical distribution of the water-rat, Nectomys spp., with schistosomiasis endemic areas, these wild rodents are very likely to acquire Schistosoma mansoni infection in their daily activities. The role of the water-rat in the S. mansoni cycle would be substantiated if one could prove that these rodents acquire the parasite during their own activity time, a completely independent time schedule of human activities. To pursue this goal, we performed two field experiments in the municipality of Sumidouro, State of Rio de Janeiro, Brazil, a schistosomiasis endemic area where N. squamipes is found naturally infected. One experiment was devised as a series of observations of activity time of the water-rat. The other experiment was a test of the occurrence of late transmission of S. mansoni to the water-rat. The daily activity pattern showed that the water-rat is active chiefly just after sunset. At both diurnal and late exposition essays the water-rat sentinels got infected by S. mansoni. These findings clarify ecological and behavioral components necessary to the adaptation of S. mansoni to the water-rat as a non human definitive host and the existence of a transmission cycle involving this animals as a reservoir.
Key words: Nectomys squamipes - Schistosoma mansoni - daily activity pattern - host reservoir The common rat, Rattus rattus, is an important element of the Schistosoma mansoni cycle in Guadaloupe focci, where three chronobiological patterns for cercarial release were observed-early, intermediate and late in daylight time _ depending on the ecological features of the transmission foccus (Théron 1984, 1985). The late pattern was specifically related to the rodent since it facilitates the encounter of the parasite with this alternative host. It was considered as a probable result of selective adaptation process (Combes & Théron 1977, Combes & Imbert-Establet 1980). Such pattern was demonstrated to be as a genetically controlled adaptive behavior (Théron & Combes 1988, Théron 1989). In Brazil, the role of the water-rat, Nectomys spp., in the transmission of S. mansoni is still controversial (Mello 1991, Chieffi et al. 1994). Due to the semi aquatic habits and the overlap of the geographical distribution of two species of Nectomys (N. squamipes and N. rattus) with schistosomiasis endemic areas, these wild rodents are very likely to acquire S. mansoni infection in their daily activities (Ribeiro et al. 1998). These widespread species have as natural habitat the moist banks of water courses (Ernest & Mares 1986), being a very common inhabitant of peri-domestic human dwellings in Brazilian rural areas (D'Andrea et al. 1999). The water-rat is known to be high susceptible to S. mansoni infection (Souza et al. 1992, D'Andrea et al. 2000), capable to eliminate viable eggs (Silva et al. 1992, Maldonado et al. 1994) and proved to be able to complete de parasite cycle (Antunes et al. 1973). These characteristics were considered as indicating Nectomys as a potential schistsomiasis reservoir in Brazil (Picot 1992, Rey 1993, D'Andrea et al. 2000). Previous studies (Rey 1993, D'Andrea & Gentile 2000, D'Andrea et al. 2000) have corroborated the hypothesis that the water-rat acts as schistosomiasis reservoirs in Brazil (Amorim 1953). This role of the water-rat in the S. mansoni cycle would be substantiated if one could prove that the rodents acquire the parasite during its own activity hours, a completely independent time schedule of human activities. To pursue this goal, we performed two field experiments in the municipality of Sumidouro, State of Rio de Janeiro, Brazil, where N. squamipes is found naturally infected. One experiment was devised as a series of observations of activity time of the water-rat. The other was a test of the occurrence of late transmission of S. mansoni to the water-rat. The epidemiological relevance of the obtained results to schistosomiasis transmission is discussed. MATERIALS AND METHODS
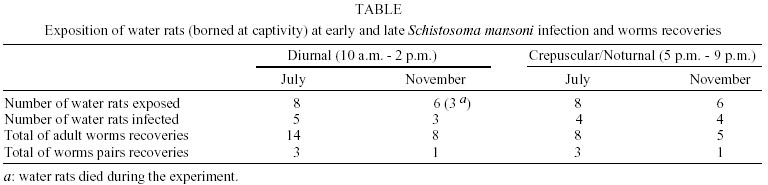
Study area - The experiments were carried out at Pamparrão and Porteira Verde adjacent valleys in municipality of Sumidouro, State of Rio de Janeiro, Brazil (22o02'46"S; 42o41'21"W). These valleys are part of a common hydric system with many small permanent streams and irrigation channels. The landscape is composed of small rural properties and few small Atlantic Forest fragments, and is characterized as low endemicity area for schistosomiasis. Previous studies in this area showed the occurrence of Biomphalaria glabrata as the intermediate S. mansoni host (Giovanelli et al. 2001) and the participation of the water-rat, N. squamipes as a definitive host for the trematoda (D'Andrea et al. 2000). The region has a humid-mesotermic climate (Nimer 1989). Rainfall is seasonal and heaviest between September and May (normal temperatures between 23.2oC and 25.4oC, normal rainfall between 153.5 mm and 269.4 mm). The driest months generally extend from June to August (normal temperatures between 18.3oC and 20.3oC, normal rainfall between 24.7 mm and 44,2 mm). The mean temperature during the study period was 24.3 ± 2.4oC and the mean rainfall was 121.7 ± 126.1 mm. Experiment 1 - Daily activity pattern of the water-rat, N. squamipes - The water-rat daily activity pattern was measured as the frequency of rodents captured per unit of time. Capture sessions were carried out for three consecutive nights during three months (January, July and October). Fifteen wired live trap (40 x 20 x 21 cm) were placed on the floor, 13 m apart from each other, kept open all day long and checked in an hourly basis from 4 p.m. to 10 p.m., and at each 3 h from 10 p.m. to 7 p.m. The trap transects were established along small streams and irrigation channels with sparse high trees, shrubs and fodder grass that characterize the typical habitat of the water-rat in the study area. Traps were baited with a mixture of peanut butter, banana, oat, bacon and a piece of manioc. All animals were released at the same station that they were captured, just after data had been recorded (i.e, individual identification mark, body mass weigh, sex, time of capture). Experiment 2 - Late infection of the water-rat by S. mansoni - We performed two essays (in July and November) using 28 N. squamipes born in captivity. At each essay, one group of rodents was exposed to S. mansoni natural infection between 5 p.m. and 9 p.m. and other group (control group) was exposed at the same location, but from 10 a.m. to 2 p.m.. The schedule of the experiment is presented in the Table. Rodents were contained individually at a perforated floating polypropylene cage that allowed cercarial penetration through skin and tail that were in contact with the water. Each cage was fixed in a station along the same streams at the same sites where the experiment 1 was carried out. After field experiment, the sentinels rodents were maintained at our laboratory during five weeks (pre-patent phase) until the necropsy. Coprologic examinations by Kato-Katz technique (Katz et al. 1972) were carried out twice a week, starting after the 4th week. After 9 weeks from exposition, the animals were killed and submitted to portal-hepatic perfusion technique (Smithers & Terry 1965) followed by collection, identification and quantification of worms from mesenteric veins. Chi-square test was used to test data between control and experimental group of the second experiment. Probability values of less than 0.05 were considered significant. RESULTS AND DISCUSSION
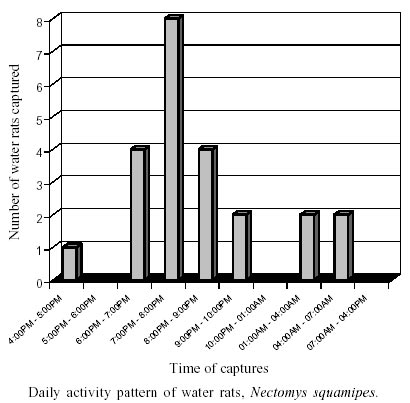
The results of time of capture observations showed that the water-rat is active chiefly just after sunset with 69.6% of captures occurring between 6 p.m. and 9 p.m. (Figure). The second experiment showed that at both diurnal and late exposition periods the water-rat sentinels got infected by S. mansoni. There was no significant difference between the number of animals infected at diurnal and late exposition in the two scenarios (c2 = 0.10, p > 0.25 and c2 = 0.14, p > 0.25, respectively), neither between total worm load (c2 = 2.3, p > 0.75) (Table). These results suggested similar risk of transmission between diurnal and nocturnal exposures. All infected water-rat showed to be able to maintain the S. mansoni cycle, eliminating viable miracidia that successfully infected snails that released cercariaes. From 16 infected animals, 56.2% individuals showed S. mansoni unisexual infection, probably due to the short period of exposition. The overall sex ratio of worms was 3.37 bias towards males (4.45M:1F at diurnal period and 2.25M:1F at late period). The same pattern was observed in natural condition for R. rattus in Guadaloupe (Théron et al. 1992, Barral et al. 1996) and for N. squamipes in a previous study at Sumidouro (D'Andrea et al. 2000). This bias towards male schistosomes was observed by Boissier et al. (1999) who described a higher life-cycle performance than females with respect to cercarial infectivity. The cercarial emergence pattern is a behavioral trait of major epidemiological interest to understand the trematode transmission, allowing the distinction between the cercariae that can infect human or other vertebrate hosts (Mouchet et al. 1992). It is well known that S. mansoni cercarial emergence occurs mainly in the circadian diurnal phase that is coincident to most human water contact activities. In our study area, human water contact is mainly diurnal, as it is related to young recreation and agricultural routines (MS Soares, in litt.). However, the S. mansoni cercarial peak shedding is heterogeneous during the day (Théron & Pointier 1995) revealing a large polymorphism in this chronobiological phenomena. This variation on the acrophase of cercarial emergence was related to the adaptation to different animal host cycles (Combes et al. 1994). Nocturnal cercarial emergence in the B. glabrata/S. mansoni system (6 p.m. to 9 p.m.) was also described within laboratory conditions (Bógea et al. 1996). In Sumidouro, Machado-Silva (1981) found two peaks of cercarial emergence from B. glabrata naturally infected: 11 a.m. and 3 p.m. This latter pattern may be related with the most activity pattern of the water-rat described herein. These findings, together with our observations on the rodent daily activity pattern, suggest the existence of a transmission cycle adapted to the participation of the water-rat as a host-reservoir for S. mansoni. These results compound a scenario in which the water-rat may act as a sort of biological filter selecting clones of S. mansoni cercariaes originated from late shedding pattern. In a population dynamic study of the water-rat, using capture-mark-release method at Sumidouro (Gentile et al. 2000), a pattern of short movement and very low migration rates for N. squamipes was detected. Maroja (2001), studying the genetic structure of the same population, found that "there was a significant allelic and genotypic differentiation among different site capture populations, and despite they were separated by very small distances, they were not panmictic, and migration was probably restricted". Such rodent sedentary behavior should have implications on rodent host-parasite relationship contributing to local parasite adaptation process. Sire et al. (2001), demonstrated clear genetic differentiation between S. mansoni sylvatic populations from a limited endemic area, and related this isolation to low gene flow due to fragmented landscape of patchy water habitats and a marked territoriality and short individual movements of the R. rattus, the only definitive host in that study area. The results found here clarify ecological and behavioral components necessary to the process of adaptation of the S. mansoni to N. squamipes as a non-human definitive host, being able to maintain the parasite cycle in nature.
ACKNOWLEDGEMENTS
To CEV Grelle and the people from the LBCE/Fiocruz for helping on the field work; to LS Maroja, O Fernandes, R Gentile, RS Coura and JF Vilela for the English review and comments; to A Maldonado Jr. for development of the experimental perforated floating polypropylene cage for Nectomys infection and JS Garcia for technical support. To Instituto Brasileiro do Meio Ambiente e Recursoso Naturais Renováveis for the collecting license. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02178t1.jpg] [oc02178f1.jpg] |
| |||||||||

{kind=link}
{kind=link}