 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(Suppl. I) October 2002, pp. 47-52 Polymerase Chain Reaction and Restriction Fragment Length Polymorphism of Cytocrome Oxidase Subunit I Used for Differentiation of Brazilian Biomphalaria Species Intermediate Host of Schistosoma mansoni Teofânia HDA Vidigal/*, Lângia C Montresor, Andrew JG Simpson**, Omar S Carvalho*/+
Departamento de Zoologia, ICB, Universidade
Federal de Minas Gerais, Belo Horizonte, MG, Brasil *Laboratório de Helmintoses
Intestinais, Centro de Pesquisas René Rachou-Fiocruz, Av.
Augusto de Lima 1715, Belo Horizonte, MG, Brasil **Instituto Ludwig de Pesquisa
sobre o Câncer, São Paulo, SP, Brasil Work partially supported by Fapemig and Pibic/Fiocruz. Received 18 June 2002 Code Number: oc02186
The intermediate hosts of Schistosoma mansoni, in Brazil, Biomphalaria glabrata, B. tenagophila and B. straminea, were identified by restriction fragment length polymorphism analysis of the mitochondrial gene cytochrome oxidase I (COI). We performed digestions with two enzymes (AluI and RsaI), previously selected, based on sequences available in Genbank. The profiles obtained with RsaI showed to be the most informative once they were polymorphic patterns, corroborating with much morphological data. In addition, we performed COI digestion of B. straminea snails from Uruguay and Argentina.
Key words: Biomphalaria - polymerase chain reaction - restriction fragment length polymorphism - cytochrome oxidase I The development of an easy and reliable method for identification of species of Biomphalaria, intermediate hosts of Schistosoma mansoni, remains an important goal regarding schistosomiasis control efforts. Recently, different DNA regions have been proposed to be used in molecular studies as an additional tool to the morphological characterization of Biomphalaria snails. These techniques include procedures such as analysis of the region 18S of rRNA through low stringenecy polymerase chain reaction (LS-PCR) (Vidigal et al. 1996, Pires et al. 1997), polymerase chain reaction and restriction fragment length analysis (PCR-RFLP) of the internal transcribed spacer region of the DNA (ITSr-DNA) (Vidigal et al. 1998, 2000, 2001, Caldeira, 1998, 2000, Spatz et al. 1999, 2000). However, high intraespecific polymorphisms and complex profiles, produced by such methodologies, may restrict their use. Although Biomphalaria snails show a high genetic variability (Knight et al. 1991, Vidigal et al. 1994), these molecular tools have important practical implications mainly concerning aspects on, extensive intraspecific variation at the morphological level (Paraense & Deslandes 1955, Paraense 1975), small size of some specimens, and inadequate fixation of snails, which make the morphlogical study rather difficult and also complicate their correct identification. Vidigal et al. (1998), showed that PCR-RFLP analysis of the ITS region is an important approach for identifying several populations of the intermediate hosts of S. mansoni in Brazil: B. glabrata, B. tenagophila and B. straminea. Interestingly, B. straminea, from Porto Alegre, showed distinct profiles from B. straminea, from other Brazilian localities, Argentina and Uruguay. Despite its polymporhic profile, it was possible to identify two common bands (namely species-specific bands) to all populations under study. In addition, B. tenagophila, from Argentina, showed a different restriction pattern when compared with those from Brazil and Paraguay, although all populations had presented at least a band in common. Thus, the ITS region showed to be useful for distinction of Brazilian S. mansoni intermediate hosts, in spite of intraspecific variations. Cytochrome oxidase I (COI) is a mitocondrial (mt) gene with an evolution rate smaller than that observed for mtDNA and its amino acid sequence is highly conserved across phyla. Molecular tools, using mtDNA COI region, have been used in several genetic studies (Lovette et al. 1999, Matsumoto & Hayami 2000, Kurabayasshi & Ueshima 2000, Campbell et al. 2000). Recently, Clark et al. (2001) used PCR-RFLP of a portion of the mitochondrial COI as a diagnostic marker for identifying several species of Coleoptera. In snails, COI region has been used for phylogenetic analysis and for interspecific comparison within the genus Bulinus, Biomphalaria and other gastropods (Stothard & Rollinson 1997, Davis et al. 1998, Campbel et al. 2000). Campbel et al. (2000) and Jones et al. (2001), analysing Biomphalaria and Bulinus, respectively, used the ITS1-rDNA sequence regions and part of COI, and showed similar results generated by each region. Stothard et al. (2001) mentioned the use of PCR-RFLP of one COI region for the differentiation of Bulinus snails. In order to assess interspecific and intraspecific variations among B. glabrata, B. straminea and B. tenagophila and verify the possibility of their identification using another DNA region (besides ITS), we amplified a COI region and then a restriction site analysis (PCR-RFLP). MATERIALS AND METHODS
Snails and DNA extraction - B. glabrata, B. tena-gophila and B. straminea populations, from different Brazilian geographical regions (Table), were used in this study. We also included, for comparison, populations of B. straminea from several Argentinean and Uruguayan localities. Snails were morphologically identified and their DNA was extracted. Morphological characterization to identify the specimens was carried out according to Paraense (1975, 1988). Their total DNA was extracted from the foot, as described by Vidigal et al. (2000). Production and evaluation of the COI RFLP profiles - One COI fragment of the three Biomphalaria species was amplified with an universal primer described by Folmer et al. (1994) (LCO- 5'-ggTCAACAAATCATAAAg ATATTgg-3'/HCO-5'-TAAACTTCAgggTgACCAAAA AATCA-3'). PCR was carried out according to Vidigal et al. (2000) except for initial denaturing step, which was 95°C for 3 min followed by 25 cycles at 95°C for 1 min; 50°C for 1 min; 72°C for 1 min 30 sec; and a final extension step at 72°C for 7 min. PCR products were digested with RsaI and AluI enzymes [that were selected based on the sequences published by Campbell et al (2000), available in Genbank], and visualized in 6% silver-stained polyacrylamide gel. The gels were photographed with a MAVICA digital camera (Sony). Many individuals of each species according to snails distribution, in Brazil, were submitted to restriction enzyme digestion. RESULTS
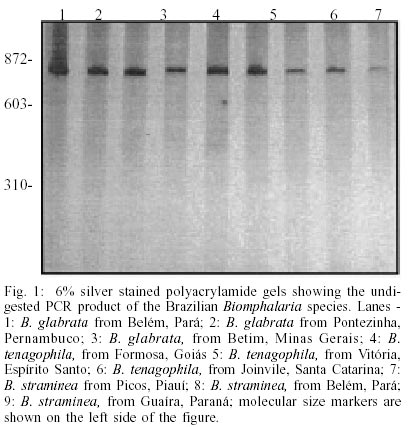
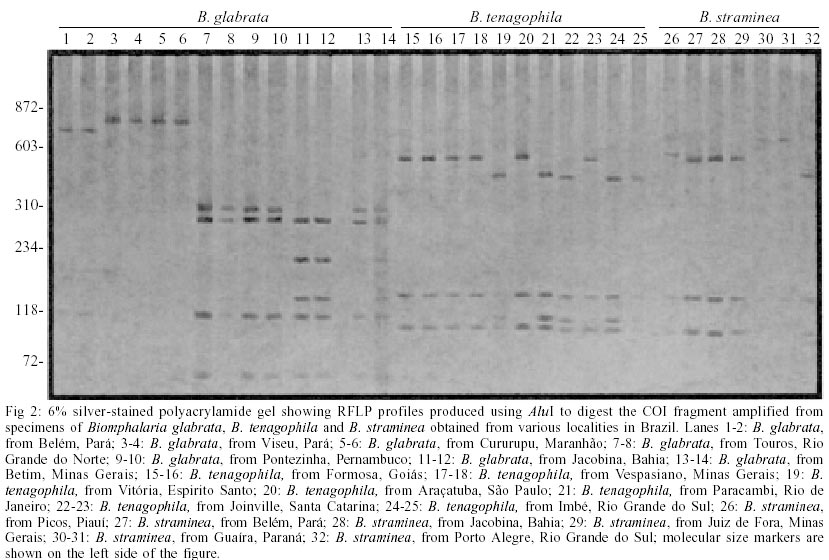
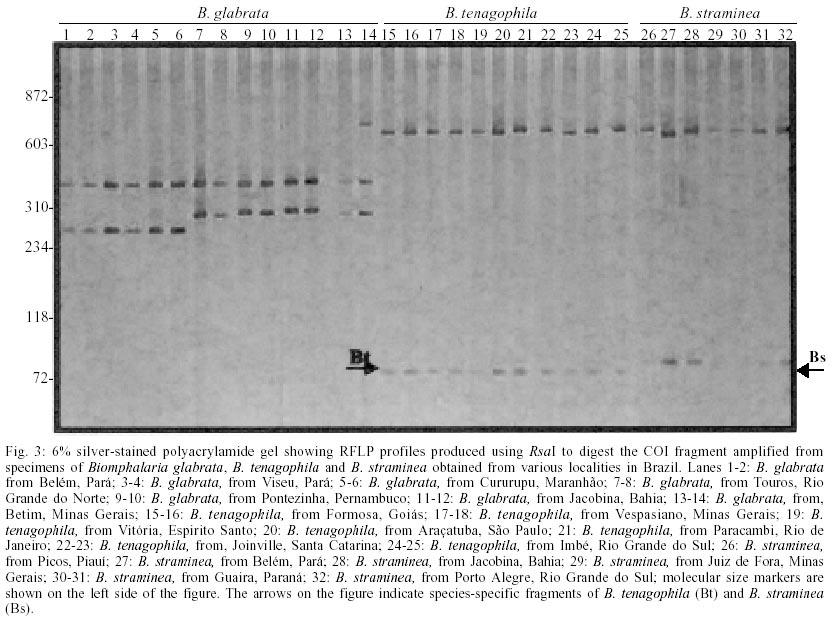
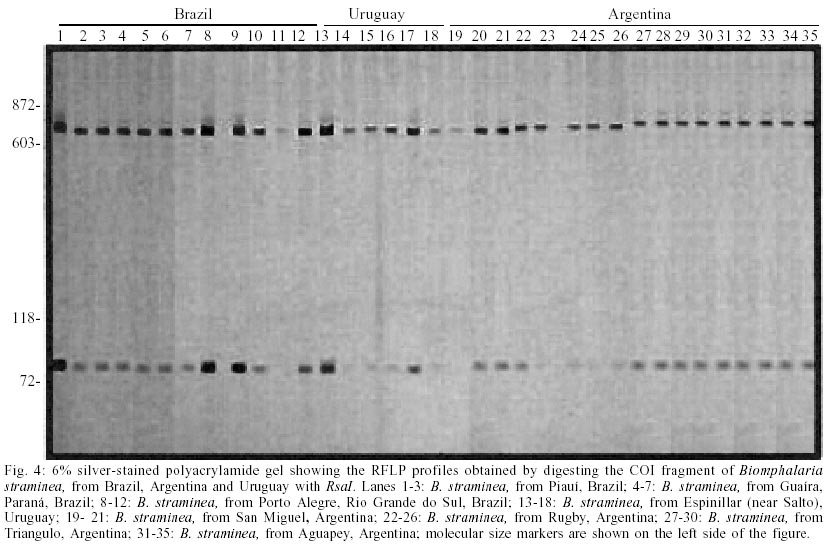
For COI region specific PCR, we obtained a single amplification product of approximately 780 base pairs (bp) for all species under study (Fig. 1). Digestion of this fragment with AluI produced highly polymorphic intraspecific and interespecific restriction profiles (Fig. 2) when we analyzed 9 populations of B. glabrata (18 individuals), 5 of B. straminea (8 individuals) and 6 of B. tenagophila (14 individuals). The best results were obtained with RsaI. For B. glabrata, we found one specific fragment (approximately 400 bp) that is present in all tested populations, however, the intraspecific profiles were not identical. Thus, B. glabrata snails were characterized by two fragments, one of 400 bp and another of 250 or 280 bp approximately, depending on the geographical origin (Fig. 3). B. tenagophila and B. straminea showed to have a similar restriction profile with two fragments (Fig. 3). The largest fragment (approximately 700 bp) did not enable the species differentiation, but the pattern of the smallest fragment (70 bp for B. tenagophila and 80 bp for B. straminea) is distinguishable. In addition, we performed COI digestion with RsaI of B. straminea snails from Brazil, Uruguay and Argentina, and obtained the same restriction band patterns of 700 and 80 bp for all individuals. DISCUSSION
Analysis of a single locus can result in incorrect species identification or lead to ambiguous conclusions (Anderson 2001). Through a comparison among results obtained with mitochondrial DNA those with ITS Blouin (2002) stated that the latter is more indicated for nematode diagnosis. Such fates suggest that it is necessary to use different regions for specific characterization, in order to achieve more reliable results. We successfully used a method based on the amplification of part of mtDNA-COI region, followed by its digestion with restriction enzymes. Inasmuch as the advantages of mithocondrial DNA applicability on sistematics and phylogenetic studies for several organisms, including snails (Casiraghi et al. 2001, Clark et al 2001, DeJong et al. 2001), are considerable. The product generated through specific PCR using the primers described by Folmer (1994) are of approximately 780 bp, value that is similar to the obtained by this author (710bp) for 11 invertebrate phyla (including Mollusca). The analysis of generated fragments (RFLP), aimed at obtaining bands to enable specific identification of B. glabrata, B. tenagohila and B. straminea, was also carried out. We performed digestions using two previously selected enzymes (AluI and RsaI). AluI digestion (Fig. 2) produced highly polymorphic intraspecific and interespecific restriction profiles indicating that this enzyme does not enable the differentiation of the three species and that there is a wide intraspecific COI variability among populations from diverse Brazilian regions. Indeed, this enzyme produced at least 4 profiles for B. glabrata, and some samples did not show restriction site for such enzyme (Fig. 2 lanes 3 to 6). Concerning B. tenagophila and B. straminea, the profiles showed to be polymorphic, although two populations of both species presented similar profiles (Fig. 2. lanes 15 to 18; lanes 26 to 29). This information agrees with those by Wilding et al. (1999) who emphasize a high polymorphism rate in mollusks mitochondrial genome. The most informative results, according to morphological data, were provided by RsaI digestion which showed to be potentially useful in these snails characterization. All enzimatic digestion of B. glabrata individuals resulted in two profiles when snails from different populations, from Brazil, were analysed. For example, the 250 bp fragment was present in populations from Maranhão and Pará, while the 280 bp fragment was present in those samples from Rio Grande do Norte, Pernambuco, Bahia and Minas Gerais. For B. straminea, we found this method to be reliable in clustering the individuals from Porto Alegre (Brazil), Argentina and Uruguay together with the populations from other Brazilian localities, as they showed the same profile (Fig. 4), different from that based on PCR-RFLP do ITS (Vidigal et al. 1998). This technique applied to COI region showed to be very effective in grouping the B. straminea populations under study (including those from Argentina and Uruguay). Despite the similarity among the size of the differential fragment, which illustrates the difference between B. tenagophila and B. straminea, these results are potentially useful for molecular identification. These results demonstrate that PCR-RFLP of this COI region, using RsaI enzyme, is an important tool for differentiation of B. glabrata, B. straminea and B. tenagophila species and they are also in accordance with those produced with PCR-RFLP of ITS, with DdeI enzyme by Vidigal et al. (1998), corroborating the Biomphalaria snails classical morphological identification. However, for the species B. straminea, PCR-RFLP of COI region showed more satisfactory results than those of the ITS region, as a single profile was produced for all samples in this trial (Brazil, Argentina, Uruguay). The current data emphasize the importance of more than one DNA region for molecular analysis. Further analyses on B. tenagophila, through the sequencing of the COI region, will be carried out for specific identification and phylo-genetic studies of Brazilian snails of the genus Biomphalaria.
ACKNOWLEDGEMENTS
To Dr Roberta Lima Caldeira from Centro de Pesquias René Rachou, for providing valuable critical comments. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02186f3.jpg] [oc02186f2.jpg] [oc02186f4.jpg] [oc02186f1.jpg] [oc02186t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}