 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(Suppl. I) October 2002, pp. 85-90 Cis-acting Elements, CArG-, E-, CCAAT- and TATA-boxes May Be Involved in Sexually Regulated Gene Transcription in Schistosoma mansoni SU Busek, M Fantappie*, LC Malaquias**, RA Wilson***, R Corrêa-Oliveira, GC Oliveira/****/+
Centro de Pesquisas René Rachou-Fiocruz,
Av. Augusto de Lima 1715, 30190-002 Belo Horizonte, MG, Brasil *Departamento
de Bioquímica, Universidade Federal do Rio de Janeiro, Rio de Janeiro,
RJ, Brasil **Laboratório de Imunologia, Universidade do Vale do Rio Doce,
Governador Valadares, MG, Brasil ***Department of Biology, University of York,
York, UK ****Programa de Pós-Graduação e Pesquisa, Santa
Casa de Misericórdia de Belo Horizonte, Belo Horizonte, MG, Brasil This study was supported by Fiocruz, CNPq, The British Council and the European Community. Received 18 June 2002 Code Number: oc02192
Schistosomes undergo various morphological and metabolic changes during their development, reflected in a finely tuned regulation of protein and/or gene expression. The mechanisms involved in the control of gene expression during the development of the parasite are not understood. Two actin genes had been previously cloned and observed to be differentially expressed during the maturation of the parasite. The SmAct gene contains four putative cis-regulatory elements (TATA-, CCAAT-, E- and CArG-boxes). Our objective was to investigate in greater detail the expression pattern of two actin genes and verify if the binding of nuclear proteins to the promoter elements of SmAct correlated with the expression profile observed. We detected little variation in the expression of actin genes during the first seven days of schistosomula culture in vitro. However, we observed significantly higher levels of expression in males compared to female adults. CArG and CCAAT elements bound to a greater extent and formed distinct complexes with male in comparison to female nuclear extracts. In contrast, female extracts bound weakly to the E-box probe while no binding was observed with male extracts. Taken together these results describe cis-acting elements that appear to be involved in sexually regulated gene expression in Schistosoma mansoni.
Key words: Schistosoma mansoni - transcription - promoter - nuclear extract - gel shift analysis During its life cycle, Schistosoma mansoni undergoes a large number of morphological and metabolic changes, dictated mainly by the constraints imposed by the radically different environments associated with each developmental stage. Schistosomes develop from a free-living water borne larval stage (cercariae) to adult worms completely adapted to the saline host environment. After penetration through the host's skin, the cercariae transform into schistosomula that become adjusted to the prevailing conditions in the mammalian host. As the schistosomula evolve to the adult stage, the parasites display marked sexual dimorphism, which implies physiological peculiarities associated with gametogenesis and reproduction. To a certain extent, the morphology and physiology of the mature parasites depends on their continuous interaction as a pair. A proportion of the eggs laid by the female worms are excreted from the definitive host and shed into fresh water where they hatch and release free living larvae, the miracidium, that will infect the snail, its intermediary host. In the snail, several generations occur, culminating with the release of cercariae. Clearly, adaptations to these major changes in environment must occur and are the result of finely tuned mechanisms that are reflected at the level of protein and/or gene expression. Actin, among other genes, has been observed to have its expression modulated. Two reports have previously shown, by northern analysis, that two distinct transcripts for actin were highly expressed in male in contrast to female worms (Davis et al. 1985, Skelly et al. 1993). In addition, lower levels of actin mRNA were observed in eggs, cercariae and schistosomula. Two of the actin genes have been previously cloned, and one of the clones, SmAct, contained four putative cis-acting elements (E-, CArG-, CCAAT-, and TATA-boxes) that are likely to be involved in gene transcription regulation (Oliveira & Kemp 1995). In this manuscript we seek to further investigate the pattern of expression of the two actin clones, SmAct and SmAct-2, and to verify if the identified cis-acting factors observed in SmAct show differential binding to nuclear extracts that parallels the observed expression pattern. MATERIALS AND METHODS
Parasites - Parasites were obtained from a life cycle of the LE strain of S. mansoni maintained at the Centro de Pesquisas René Rachou-Fiocruz. Schistosomula were mechanically transformed using the method previously described by Ramalho-Pinto et al. (1974) and modified by Michalick et al. (1979). Transformed schistosomula were cultivated according to previously published procedures (Michalick et al. 1979, Basch 1981). Male and female adult worms were physically separated after perfusion of infected mice as previously described (Gazzinelli et al. 1983). Clones - The clone encoding SmCOII-ND6 (accession number AF153976) was obtained by immunoscreening an adult worm expression library, and sequenced on both strands as previously described (Oliveira & Kemp 1995). Both actin clones had been previously characterized (Oliveira & Kemp 1995). RT-PCR - Total RNA was extracted using Tri Reagent (Sigma, St. Louis, USA) according to the supplier's protocol (Chomczynski 1993). For cDNA synthesis, 2.5 µg of RNA, Superscript II RNAase H-reverse transcriptase (Life Technologies, Gaithersburg, MD, USA) and an oligo-dT primer were used, following the supplier's protocol. PCR amplification was made semi-quantitative by carefully controlling the starting amount of RNA, MgCl2 concentration, annealing temperature and cycle number. The specific conditions for each primer pair are described in the Table. For all of the primer pairs, 1 µl of cDNA was added to a mixture containing the supplied reaction buffer, 10 mM of each dNTP, the appropriate amount of MgCl2 (Table), 0.5 units of Taq DNA polymerase (Life Technologies), and 10 pmol of each primer. The mixture was incubated at 95°C for 2 min, followed by the appropriate number of cycles (Table) in a thermocycler (MJR PTC-100, Watertown, USA) as follows: 95ºC for 1 min, the optimized annealing temperature for 1 min (Table), and 72ºC for 1 min, followed by a final 10 min incubation at 72ºC. The PCR products were separated by electrophoresis in a 6% polyacrylamide gel in TBE buffer using a Mini-Protean II apparatus (BioRad, Hercules, USA). The gel was stained with ethidium bromide (Sigma) and the amplified DNA visualized using the Eagle Eye II digital imager (Stratagene, La Jolla, USA). Densitometry analysis was carried out on the digitized image using the ZeroDScan software (Stratagene). Densitometry results were normalized for the densities obtained with 0 h schistosomula in 0 to 24 h culture experiments, and for 0 to 7 days of culture experiments, which were given the arbitrary value of 100. All of the experiments were carried out in triplicate. Northern blots - Northern blot analysis was performed according to common procedures with the use of Rapid Hybridization buffer (AmershamPharmacia, Uppsala, Sweeden). For Northern analysis 5 µg of S. mansoni poly(A+) RNA, extracted with TriReagent (Sigma, St. Louis), according to the manufacturer's instructions, were fractionated in a 1.2% agarose gel in the presence of 2.2 M formaldehyde, with 0.24 to 9.5 kbp molecular weight markers (Life Technologies) and transferred to a Nylon membrane (Hybond, AmershamPharmacia). Hybridization was carried out at 65°C for 2-3 h in with approximately 2 x 106 c.p.m./ml of radioactively labeled DNA probe (Multiprime kit, AmershamPharmacia) in Rapid Hybridization buffer, according to the manufacturer's instructions. Membranes were exposed to XAR5 film (Kodak, Rochester, NY) with a hyperscreen intensifying screen (AmershamPharmacia) at -70°C for 1 day. RNase protection assays (RPA) - RPA were performed according to instructions in the MAXIscript and RPA II kits (Ambion, Austin, TX). The protected samples were loaded into the gel apparatus (VAGE, Stratagene) and run in a 5% SDS-PAGE-urea at 300 V. The gel was removed, overlaid with Whatman paper, wrapped in plastic wrap, and exposed to film or a phosphoimaging plate (Fuji, Japan). In all of the RPA experiments, Yeast RNA was used as digested and undigested controls. Quantification of the bands observed was performed by computer aided densitometry analysis of the gel image captured by the phosphoimaging equipment (phosphoimager using BAS and MacBAS software Fuji Photo Film Co.), were normalized for the highest density band, which in all experiments was the RNA sample of adult male worms. Background measurements were obtained from the region in the digested yeast RNA lane, where a band of the size of the protected fragment would be found. For comparison, a region of the image where no bands were expected was also used as a background measurement. The relative results obtained were essentially the same no matter what background measurement was taken. Band shift assay - End-labeled DNA (50-100.000 cpm ~ 50ng) was incubated with male and female nuclear extracts (Elton & Reeves 1985) for 15 min at room temperature, in a final volume of 30 µl. The binding buffer contained 10 mM Tris-HCl, pH 7.5, 50 mM NaCl, 0.5 mM DTT in 10% glycerol, plus 1mg of acetylated bovine albumin and 1 µg of poly-d(I)d(C) (AmershamPharmacia) used as a competitor DNA to block non-specific protein-DNA interactions and 20 fmol of P32 labeled DNA. After incubation, the samples were fractionated in a 4% non-denaturing polyacrylamide gel using 0.5x TBE buffer at 120 V for 3 h at 4°C. The gel was dried under vacuum and exposed to an X-ray film with an intensifying screen. The double stranded DNA probe sequences (Life Technologies) used in these experiments were as follows, 5' to 3': CCAAT-box TAAAGTCATAAATGTAGA, TATA-box GTATTT AGTAAAGTGAAT, E-box TGAATCAAGTGTCTCA, CArG box GTTAGCCATTATGGCATT. Statistical analysis - All statistical analysis was conducted using ANOVA at a 95% confidence level. RESULTS
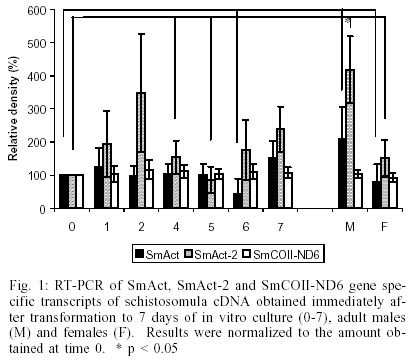
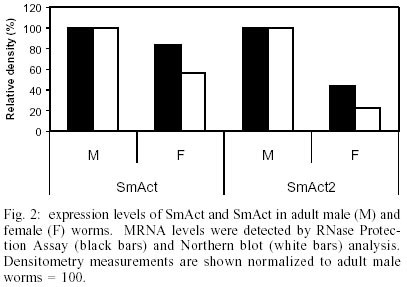
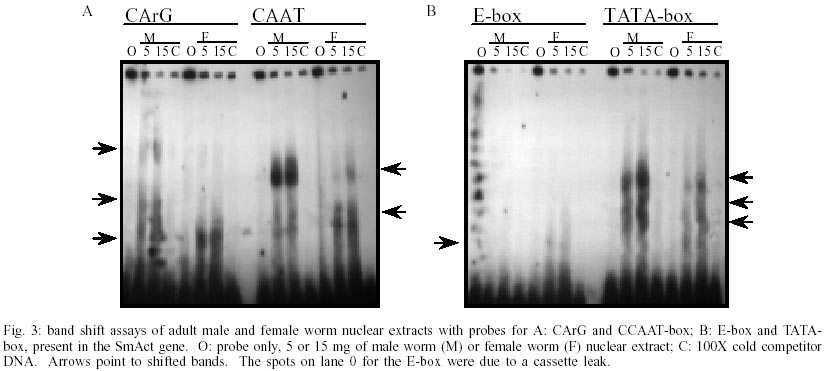
The transcription levels of the three genes, SmAct, SmAct-2 and SMCOII-ND6, were initially investigated during the first 24 h after mechanical transformation of cercariae to schistosomula. We did not observe any statistically significant difference in transcript levels as detected by RT-PCR (data not shown). The only exception was a slight increase in SmAct transcript levels at 8 h after transformation (p = 0.002). We followed the initial experiments for 24 h after transformation with the observation of the transcript levels for the next seven days of in vitro culture, in 24 h intervals. In Fig. 1 we can observe that there is a small fluctuation in the levels of SmAct and SmAct2 transcript levels during the first 24 h period, detected by RT-PCR, with a greater variation for the latter gene. SmAct transcript levels at day 6 were significantly lower from what was observed for mature male worms. SmAct-2 transcripts at days 0, 4 and 5 were significantly lower when compared to mature male worms. For the same period we did not observe any significant fluctuation in the levels of SMCOII-ND6 transcripts. Compared to day 0 of culture, slightly higher levels of SmAct, and considerably higher levels for SmAct2 transcripts were observed in adult males. Both genes showed higher levels of expression in adult males when compared to females. By RPA and northern blot analysis, male adult worms displayed higher levels of SmAct and SmAct2 transcripts than female worms (Fig. 2). Again, the levels of SMCOII-ND6 transcript did not vary between sexes. We also investigated the differences in transcription levels between whole cercariae, cercarial tails and eggs. We observed that the levels of transcripts were not significantly different for any of the three genes, with the exception of somewhat lower transcript levels of SmAct-2 in cercarial tails (data not shown). By RPA and northern blot, the transcript levels of all three genes were lower in cercariae than in adult worms (data not shown). The observation of different levels of transcript for SmAct and SmAct-2, especially between sexes, led us to investigate whether the putative cis-acting transcription regulatory elements, the two ubiquitous TATA- and CCAAT-boxes and the more restricted E- and CArG-boxes would show different binding to nuclear extracts of male or female adult worms. We did not test the other life cycle stages due to the difficulty in obtaining large enough quantities of the extracts. In Fig. 3A, it can be observed that CArG-box derived probes bound to nuclear extracts of male and female worms. Male worm-derived nuclear extracts bound to a greater extent to the CArG probe then did female extracts. Interestingly, two of the higher molecular weight complexes formed with the CArG probe and male adult worm extracts were not observed for female nuclear extracts. Fig. 3B shows that E-box elements bound weakly to female worm nuclear extracts. No binding was observed for male worm nuclear extracts. TATA- and CCAAT-box derived probes bound to male and female nuclear protein extracts, but the level of binding was greater for the former (Fig. 3A, B). For Fig. 3A, B the binding was dependent on the concentration of nuclear proteins used (0, 5 or 15 µg). Binding specificity was demonstrated by competition of excess unlabeled DNA (100X). DISCUSSION
In this paper we describe by three different methods, RT-PCR, northern blot and RNase protection assay, that two actin gene transcripts are expressed at higher levels in adult male worms than females. In addition, we describe cis-acting elements (CArG-, CCAAT-, E-box) that may be involved in sexually regulated transcription of genes. Other workers using a heterologous (Davis et al. 1985) or a homologous probe (Skelly et al. 1993) described similar levels of actin expresion. Both describe two hybridizing bands of approximately 1.4 and 1.9 kb, which probably represents the two genes we studied (Oliveira & Kemp 1995). We also demonstrate in detail that there are fluctuations of the expression levels of these two genes in the first 24 h and the first 7 days of schistosomula maturation in vitro. Cercariae and eggs also contain lower amounts of transcripts for the two genes, in accordance with previous descriptions (Skelly et al. 1993). Similar to our results, the same report did not show higher mRNA levels for actin during the first 24 h after transformation (Blanton & Licate 1992). In contrast, at the protein level, a two-dimensional electrophoresis (2-D) spot with molecular weight and PI characteristic of actin was observed at higher intensities 24 h after transformation into schistosomula (Blanton & Licate 1992). Actin was also shown to be expressed at higher levels in males in comparison to females at the protein level by 2-D electrophoresis (Atkinson & Atkinson 1982, Abbas & Cain 1989). Modulation of actin mRNA was also observed after in vitro treatment of adult worms with a schistosomicidal compound (Eshete & Bennett 1991). In addition to actin, we explored the expression levels of a mitochondrially encoded transcript containing the SmCOII/ND6 genes. We did not observe any significant variation in the mRNA levels for this transcript by any of the three techniques used. This observation agrees with the previously described stable expression of another mitochondrial transcript, SCOX1 (Skelly et al. 1993). We suggest that these two mitochondrial transcripts could be used as controls for experiments that investigate genome wide gene expression levels using e.g. microarrays. Very little is known about how schistosomes regulate gene expression. One of the most comprehensive analyses has been carried out with the regulatory domain of the Sm28 (GST) gene. Functional Y box regulatory sequences have been found in the promoter region of Sm28 and shown, in heterologous transient transfection experiments, to be responsible for most of the promoter activity (Serra et al. 1996, 1997). In addition, the female-specific gene F10 that was shown to contain regulatory elements that bound differentially to male or female protein extracts (Engelender et al. 1993, Giannini et al. 1995). CCAAT-box is an ubiquitous cis-acting element usually positioned about 50 to 100 bp upstream of the start site, and is involved in promoter function (Johnson & McKnight 1989). TATA-box is another common cis-acting element that is found associated with many genes (McKnight & Kingsbury 1982, Singer et al. 1991). It is usually located 40 to 100 bp upstream of the transcription start site and is functionally involved in fixing the start of transcription (McKnight & Kingsbury 1982, Jones et al. 1988). The binding of schistosome nuclear proteins to CCAAT elements has been observed before, with results similar to those shown here (Serra et al. 1996). CArG boxes are present in a variety of genes, from plants to several vertebrate species (Carroll et al. 1988, Tilly et al. 1998, Froman et al. 1998). CArG boxes are related to the known serum response elements (SRE), and are thought to be a part of the core machinery for muscle-specific transcription of several genes (Bergsma et al. 1986, Carroll et al. 1988, Nakamura et al. 2001). The activation of muscle-specific genes is thought to require the binding of serum response factors (SRF) (Soulez et al. 1996, Strobeck et al. 2001) in addition to transcription factors such as GATA-4, Barx1b, Nkx2.5 and phox/Mhox to the CArG box (Grueneberg et al. 1992, 1995, Belaguli et al. 2000, Nakamura et al. 2001, Chang et al. 2001). E-boxes have been found to participate in the transcriptional regulation of a large variety of genes in a cell or tissue specific manner (Hillyer et al. 1988, Sharma et al. 1996, Matsuda et al. 1997) and to respond to basic-helix-loop-helix (bHLH) transcription factors that are associated with gene regulation during cell differentiation (Chaudhary et al. 1997). MyoD is one of the bHLH trans-acting factors known to interact with the E-box element to drive transcription of muscle specific genes in many cell types (McKnight & Yamamoto 1992, Moss et al. 1994). Other myogenic transcription factors that bind to E-boxes are Myf5, myogenin and Mrf4 (Molkentin & Olson 1996, Cossu et al. 1996). The transcription of Fgf4, a signaling molecule that plays a major role in cell differentiation, has also been found to be dependent on the binding of Myf5 to an E-box present in the promoter region (Fraidenraich et al. 2000). Other factors that bind to the E-box, such as MyoR can act as a repressor of muscle specific transcription (Lu et al. 1999). The binding of female nuclear extract proteins to the E-box probe may indicate the presence of such a repressor factor, as both SmAct and SmAct-2 genes were observed to be expressed to a smaller degree in adult female worms. The knowledge obtained by investigating mechanisms of transcription control would greatly benefit efforts into expressing laboratory constructs in the parasite. The results presented here indicate that possibly transcription factors similar to the above may be found in schistosomes. The differential binding of male and female nuclear extracts to the cis-acting elements studied could be interpreted as quantitative and/or qualitative differences that may be related to sex-specific or -preferential transcription. These results can be confirmed with further cold probe competition experiments. Future experiments will aim at investigating the trans-acting factors that interact in particular with the E- and CArG-boxes. MyoD and SRF have been shown to bind to these cis-elements in other systems (McKnight & Yamamoto 1992, Moss et al. 1994, Soulez et al. 1996, Strobeck et al. 2001). None of these genes have yet been cloned in schistosomes or identified by EST sequencing. It will be interesting to investigate the role of these transcription factors in gene regulation in schistosomes. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02192t1.jpg] [oc02192f3.jpg] [oc02192f2.jpg] [oc02192f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}