 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(Suppl. I) October 2002, pp. 91-93 SHORT COMMUNICATION
Partial Molecular Characterization of Sm8, a Tegumental Antigen of Schistosoma mansoni Frederico GC Abath+, Edeneide M Xavier, Sílvia ML Montenegro, Roberto P Werkhauser
Centro de Pesquisas Aggeu Magalhães-Fiocruz,
Av. Moraes Rego s/no, Cidade Universitária, 50670-420 Recife,
PE, Brasil This work was supported by grants from Brazilian Research Council (CNPq). Received 18 June 2002 Code Number: oc02193
Sm8 is a major tegumental antigen of Schistosoma mansoni. The partial cDNA was isolated and analyzed. Sequence analysis revealed transmembrane compatible hydrophobic domains and a putative leucine zipper pattern. The mRNA and the protein are predominantly expressed in adult worms.
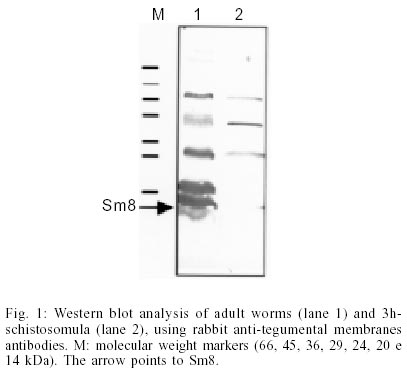
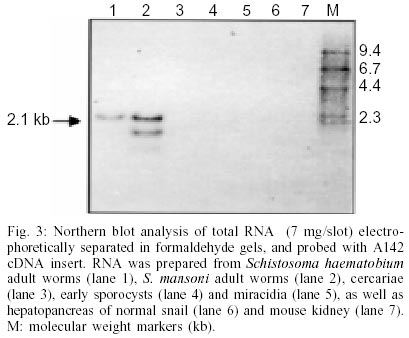
Key words: Schistosoma mansoni - tegumental antigen - characterization It has been shown that isolated Schistosoma mansoni adult worm tegumental membranes are capable of stimulating protective immunity in mice (Smithers et al. 1989, 1990). Additionally, the principal membrane associated antigens contained in adult S. mansoni tegument do not cross react with the egg antigens of the parasite which are involved in immunopathology (Smithers et al. 1990). Thus, the characterization of proteins within the tegument is relevant in a more basic level to improve the understanding of the function of this structure and in a more applicative level to identify molecules that are useful for diagnosis, or may act as targets of protective immunity and/or chemotherapy (Abath et al. 2000). Several genes for major antigens recognized by antibodies from mice protectively immunized with tegumental membranes (Smithers et al. 1990) were cloned and characterized: Sm25 (Omer-Ali et al. 1991); Sm22 (Omer-Ali et al. 1991); Sm15 (Abath et al. 1993), Sm13 (Abath et al. 2000). In the present communication, we will extend theses studies reporting on the partial characterization of an 8 kDa tegumental antigen (Sm8). Raising of antisera, screening of cDNA libraries and subcloning, DNA sequencing and analysis, Northern blot and Western blot analysis were carried out as previously described (Abath et al. 1994, 2000). Western blot analysis, using rabbit antibodies raised against tegumental membranes, showed that Sm8 is recognized in tegumental preparations of adult worms but not 3h-schistosomula (Fig. 1). The cDNA for Sm8 (A142 cDNA) was isolated by screening a lgt11 adult worm S. mansoni cDNA library with rabbit anti-adult worm tegumental membrane antisera (Abath et al. 1994).The cDNA was sequenced (Fig. 2, GenBank AF313910), and shown to contain 356 nucleotides. However, the apparent molecular mass of the mRNA as determined from Northern blot analysis is 2.1 kb (Fig. 3). As the blot was hybridized with different probes, additional bands, representing different messengers can be also seen in Fig. 3. The mRNA for Sm8 was detected in adult worms, but not in miracidia and sporocysts. This is in agreement with the pattern of protein expression, as evaluated by Western blot analysis, that showed predominance of the antigen in adult worms (Fig. 1). Interestingly, the messenger was also detected in S. haematobium adult worms, indicating a certain degree of homology. With specific regard to tegumental surface antigens, the modulation of expression during maturation may result in the disappearance of some antigens while new polypeptides may appear (Simpson et al. 1984), with profound consequences for the outcome of the immune response elicited by the Schistosoma infection. Although the sequence contains the stop codon, the codon for the initial methionine is missing, and the size of the mRNA is bigger than the isolated cDNA, indicating that the gene cloned is not complete. As yet the function of Sm8 is unknown, and the coding region contains no homologies with other proteins in the currently available data bases. Maybe this would indicate that this protein has a unique function in the tegument of S. mansoni. Searching the available EST data banks we found a homologue of the Sm8 cDNA (EST272900), that although containing additional 24 bp toward the 5'end, did not reach the initial methionine codon. Fig. 2 shows the resulting collated sequence. The analysis of the predicted protein suggests the presence of three possible N- terminal transmembrane helices (Fig. 2, aa 1-26, 27-45, and 50-72). The open reading frame of the gene was subcloned and expressed in pMal-cri. We have raised rabbit sera against the recombinant maltose binding protein fusion. However, these antibodies were not capable to recognize the parasite counterpart (data not shown), what may suggest that the recombinant protein is poorly antigenic. Indeed, the derived amino acid sequence is highly hydrophobic, particularly in the N-terminus, suggesting poor antigenicity. On the other hand, recombinant eukaryotic polypeptides synthesized in the Escherichia coli expression system, can differ from their authentic counterparts, because several eukaryotic post-translational modifications are not performed in E. coli, such as glycosylation, acetylation and amidation. The limitations of the use of eukaryotic genes expressed in bacteria in immunological studies have been previously discussed (Abath et al. 1997). The identification of Sm8 in tegumental membranes, by Western blot analysis, in conjunction with the analysis of the cDNA sequence, strongly suggest that Sm8 is a tegumental membrane protein. When searches were performed against PROSITE (ca.expasy.org), a motif compatible with a leucine zipper pattern was found (aa 30-51). This pattern has been proposed to explain how some eukaryotic gene regulatory proteins work. Nonetheless, its potential biological relevance must be validated experimentally. It is unclear how proteins that are expected to be cytosolic, such as Sm15 (Abath et al. 1993, 1994), GST (glutathione S-transferase) (Balloul et al. 1987), TPI (triose-phosphate isomerase) (Harn et al. 1992), GAPDH (glyceraldehyde-3-phosphate dehydrogenase) (Goudot-Crozel et al. 1989) and PGK (phosphoglycerate kinase) (Lee et al. 1995), could be attached to the schistosome surface, since none have a conventional transmembrane domain and there is no evidence of a lipid anchor (Abath & Werkhauser 1996). We have speculated previously that the tegumental membrane Sm13 could be interacting with at least some of these proteins allowing for the attachment to the tegumental surface (Abath et al. 2000). This speculation is also valid for Sm8, since, as Sm13, it is also coexpressed with Sm15 and other soluble protein in the tegument of hepatic worms (Xavier et al. 1998). We are currently cloning the 5'end of the cDNA and making efforts to obtain antibodies able to recognize native Sm8, and useful for immunochemical studies. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02193f1.jpg] [oc02193f2.jpg] [oc02193f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}