 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(Suppl. I) October 2002, pp. 117-127 Intestinal Fibrovascular Nodules Caused by Schistosoma mansoni Infection in Calomys callosus Rengger, 1830 (Rodentia: Cricetidae): a Model of Concomitant Fibrosis and Angiogenesis Jane A Lenzi/+, Ester M Mota, Marcelo Pelajo-Machado, Leandro S Vale, Bruno S Vale, Zilton A Andrade*, Henrique L Lenzi
Departamento de Patologia, Instituto
Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
*Centro de Pesquisas Gonçalo Moniz-Fiocruz, Salvador, BA, Brazil
This work was supported by Fiocruz and CNPq. Received 18 June 2002 Code Number: oc02200 Human schistosomiasis develops
extensive and dense fibrosis in portal space, together with congested new blood
vessels. This study demonstrates that Calomys callosus infected with Schistosoma
mansoni also develops fibrovascular lesions, which are found in intestinal subserosa.
Animals were percutaneously infected with 70 cercariae and necropsied at 42,
45, 55, 80, 90 and 160 days after infection. Intestinal sections were stained
for brightfield, polarization microscopy, confocal laser scanning, transmission
and scanning electron microscopies. Immunohistological analysis was also performed
and some nodules were aseptically collected for cell culture.
Key words: Calomys callosus - Rodentia - Cricetidae - Schistosoma mansoni - intestine - nodules - fibrosis - angiogenesis Important characteristics of the human chronic schistosomiasis mansoni in portal space pathology are extensive and dense fibrosis, together with well vascularized and often tortuous, thin-walled, dilated, and congested blood vessels, presenting a varicose or angiomatoid appearance (Bogliolo 1954, Andrade 1965). The new vessels have arterial and venous (periductal plexus) origin (Bogliolo 1957, Andrade & Cheever 1971) and the mechanism of this angiogenesis is still unknown. Otherwise, it has been assumed that hepatic fibrogenesis is related to the activity of fibroblasts, myofibroblasts and Ito cells (Grimaud & Borojevic 1977, Popper & Martin 1982). However, to our knowledge, none of the described experimental models presents the concomitant association of fibrosis and angiogenesis related to schistosomal infection (Lichtenberg et al. 1971, Cheever 1987). In this study we show that the rodent Calomys callosus infected with Schistosoma mansoni develops prominent intestinal fibrovascular nodules which show the concomitance of fibrosis and angiogenesis in periovular reaction. MATERIALS AND METHODS
C. callosus were infected when they were five days old by percutaneous exposure to 70 cercariae of the Belo Horizonte (BH) isolate of S. mansoni (Paraense & Corrêa 1963, 1981). The rodents were necropsied at 42 (20 Cc), 45 (30 Cc), 55 (10 Cc), 80 (10 Cc), 90 (30 Cc) and 160 (30 Cc) days after infection. Intestinal samples were prepared according to Swiss roll procedure (Lenzi & Lenzi 1986) and fixed in Carson's Formalin-Millonig (Carson et al. 1973) and embedded in paraffin. Sections (5 µm) for brightfield microscopy were stained with hematoxilin and eosin; PAS-Alcian Blue, pH = 1.0 and 2.5; Lennert's Giemsa, Picrosirius (plus polarization microscopy) (Junqueira et al. 1979), Gomori's Silver Reticulin and resorcin-fuchsin stain with or without oxidation with potassium peroxymonosulfate [Oxone monopersulfate compound (Sigma - 22, 803-6)]. Another set of sections was stained with Masson's trichrome, Evans blue (5 µm) (Vale et al. 1997) and phosphomolibdic acid-Picrosirius (PMA-PSR) (20 µm) (Dolber & Spach 1993) stains for confocal laser scanning microscopy (CLSM) (LSM 410, Zeiss) to discriminate eosinophils, elastic and collagen fibers. Three-dimensional reconstructions of images were done with 20 µm sections of the intestinal nodules. Sections stained with Alcian blue-Safranine (Strobel et al. 1981, Gomez et al. 1987) were applied to distinguish the mast cell subpopulations by brightfield and confocal (CLSM) microscopies. For electron microscopic examination, sections of the intestinal nodules were fixed in 2.5% glutaraldehyde buffered with 0.1 M cacodylate and postfixed in 1% osmium tetroxide, dehydrated in graded acetone series and embedded in epoxy resin. Ultra-thin sections were stained with uranyl acetate and lead citrate and observed by an EMS 10 B Zeiss electron microscope. For scanning electron microscopy (SEM), segments of intestine were fixed in Carson's Formalin-Millonig (> 24 h), desiccated in incubator (37oC, 1 h), and analysed in SEM-low vacuum mode (LEO 435-VP). Immunohistological analysis of the intestinal nodules were performed on cryostat sections or trypsinized paraffin slides by indirect immunofluorescence or indirect peroxidase labeled extravidin or streptavidin-biotin staining (ExtrAvidin-Peroxidase, Sigma E-2886; anti-mouse polyvalent immunoglobulins biotin conjugate (Sigma, B-2016); anti-rabbit immunoglobulins biotin conjugate (Sigma, B-3275). In most of the cryostat sections, after fixation with cold acetone (4oC, 10 min) we used indirect immunofluorescence, blocking of non-specific binding by 1% bovine serum albumin and 3% skimmed milk (at room temperature, 30 min). The following primary antibodies were applied: collagen I (rabbit anti-human collagen type I, Institut Pasteur de Lyon, ref. 07.90), collagen III (monoclonal mouse anti-human collagen type III, GIBCO BRL, ref. 12070-017), collagen VI (monoclonal human anti-collagen type VI, American Qualex, ref. M2190), Factor VIII-Related Antigen = FVIII-RA (rabbit anti-human F VIII-RA, DAKO, ref. A082), fibronectin (polyclonal rabbit anti-human fibronectin, DAKO, ref. A245), laminin (polyclonal rabbit anti-mouse laminin, American Qualex, ref. R1990), tenascin (polyclonal rabbit anti-human tenascin, American Qualex, ref. R2230), chondroitin-sulfate (monoclonal mouse anti-chicken chondroitin sulfate, Sigma, ref. C8035), alpha smooth muscle actin (monoclonal mouse anti-alpha smooth muscle actin, Sigma, ref. A-2547), desmin (monoclonal mouse anti-pig desmin, Sigma, ref. D-1033), vimentin (monoclonal mouse anti-pig vimentin, Sigma, ref. V-6630) and pan-cytokeratin (rabbit anti-bovine muzzle epidermal keratin, American Qualex, ref. R0760). Goat anti-rabbit IgG conjugated to rhodamine (American Qualex, A102RN) and goat anti-mouse IgG + IgM + IgA conjugated to rhodamine (American Qualex, A157RN) were used as secondary antibodies. Immunofluorescence-stained samples were analyzed with an LSM 410 Confocal Laser Scanning Microscope (Zeiss, Germany). Digital images of fluorescence were acquired in response to excitation with a Helium/Neon laser (543 nm) (filter setting BP 575-640). Nodules of three C. callosus with 90 days of infection were aseptically collected by mechanical dissection in laminar flow after death of the animals by ether. Isolated nodules were cultivated in six-wells plate, in McCoy's 5A medium (Sigma - M4892), supplemented with 20% fetal bovine serum and penicillin-streptomycin (10 µl/ ml of total medium) in 5% CO2 incubator, 37oC. RESULTS
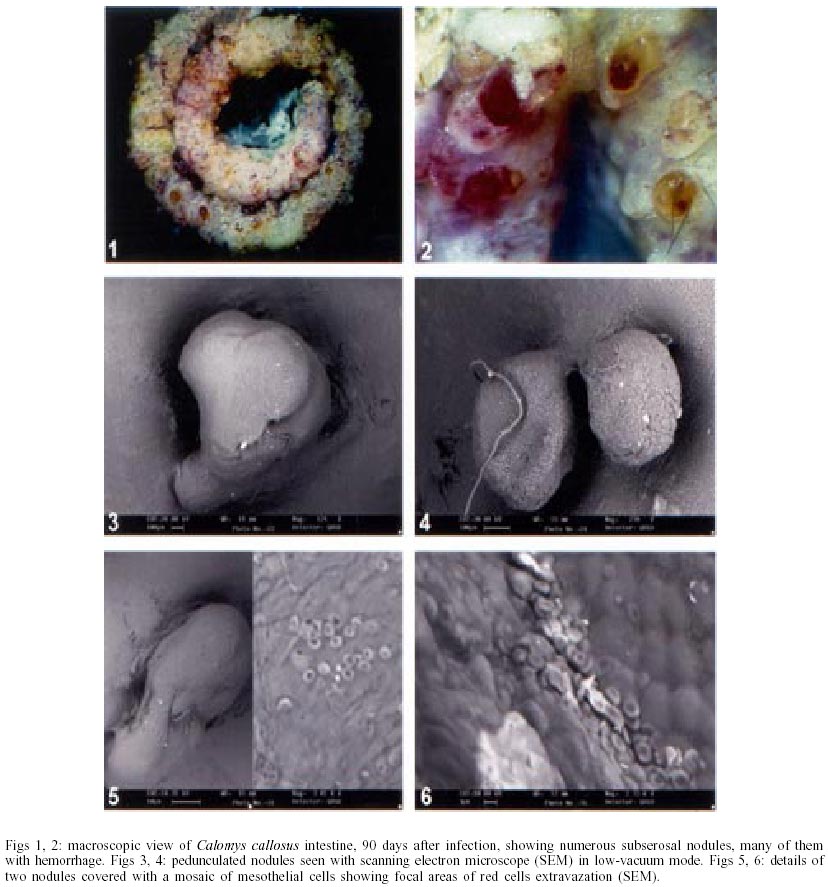
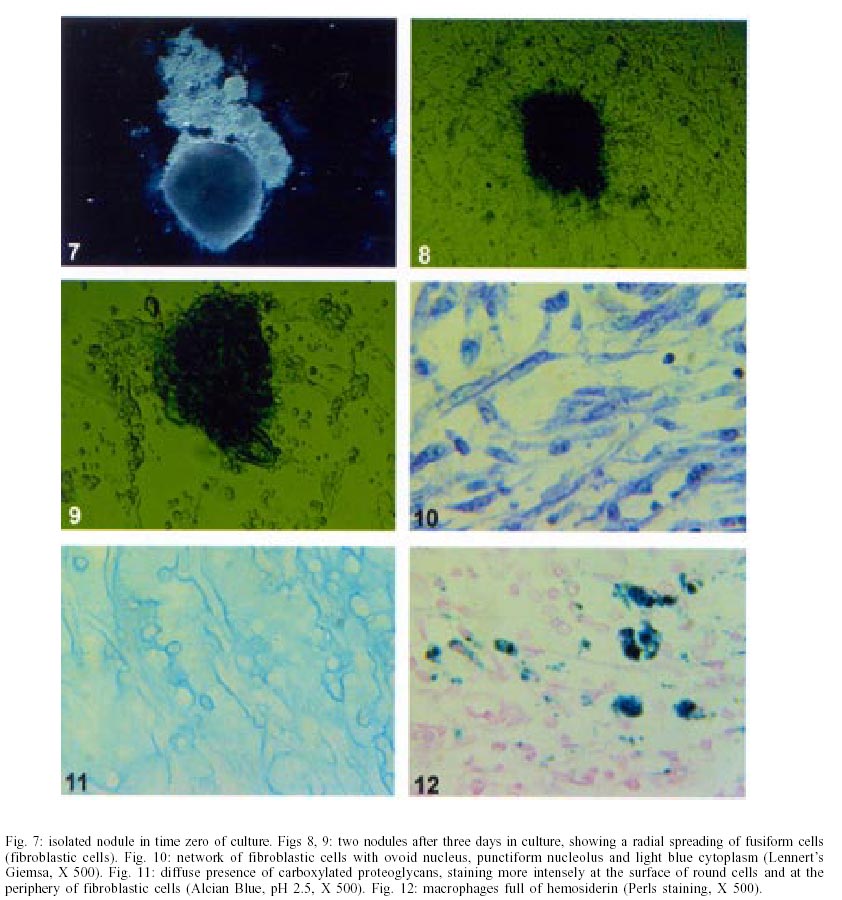
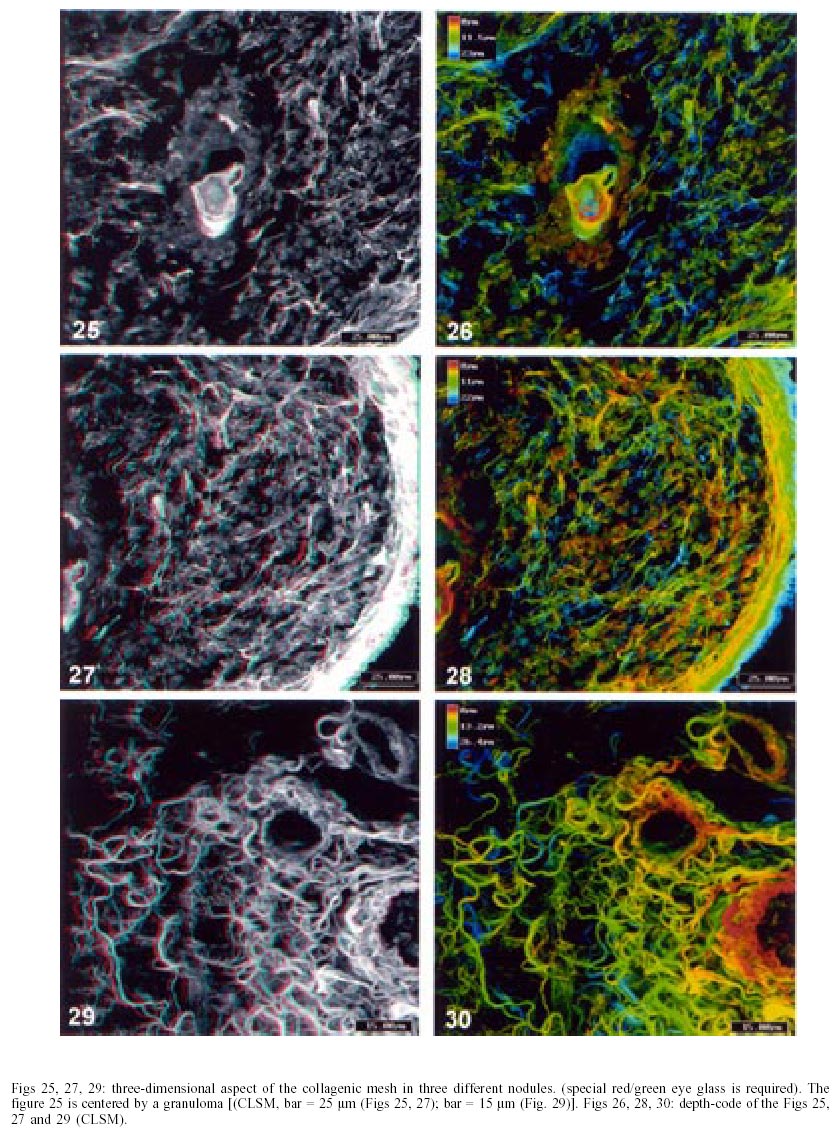
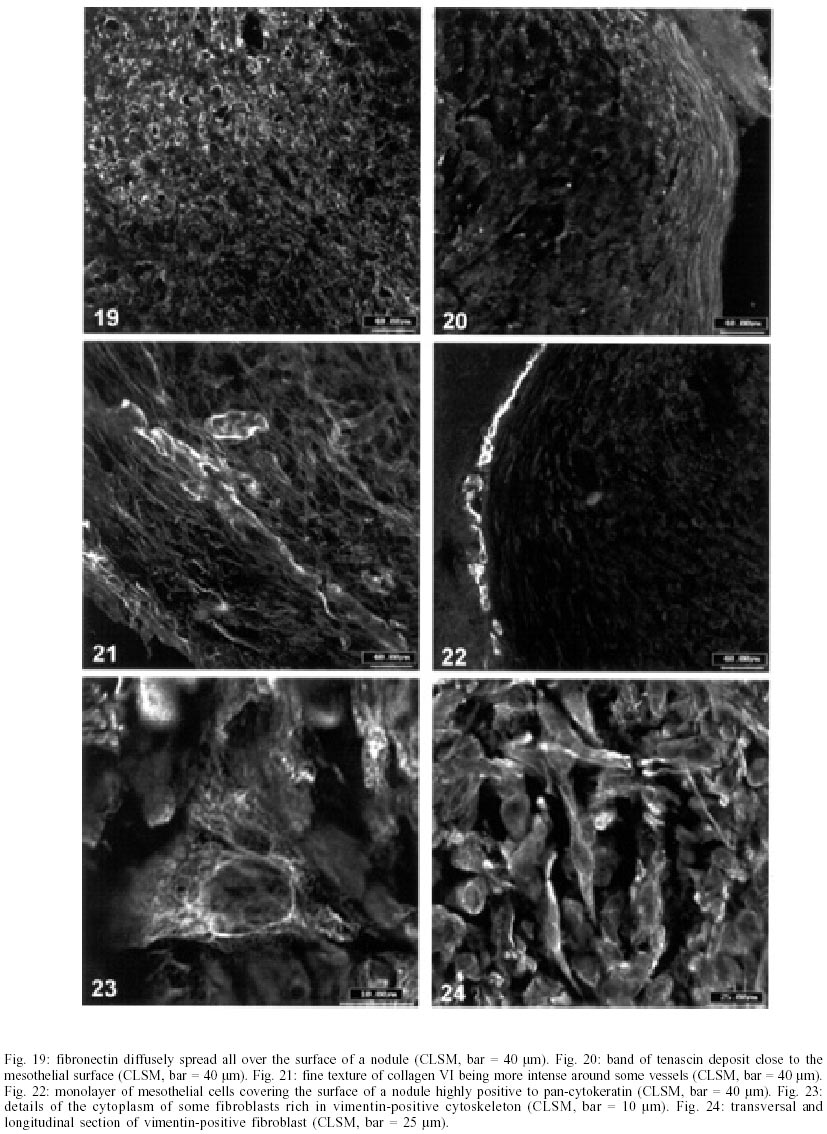
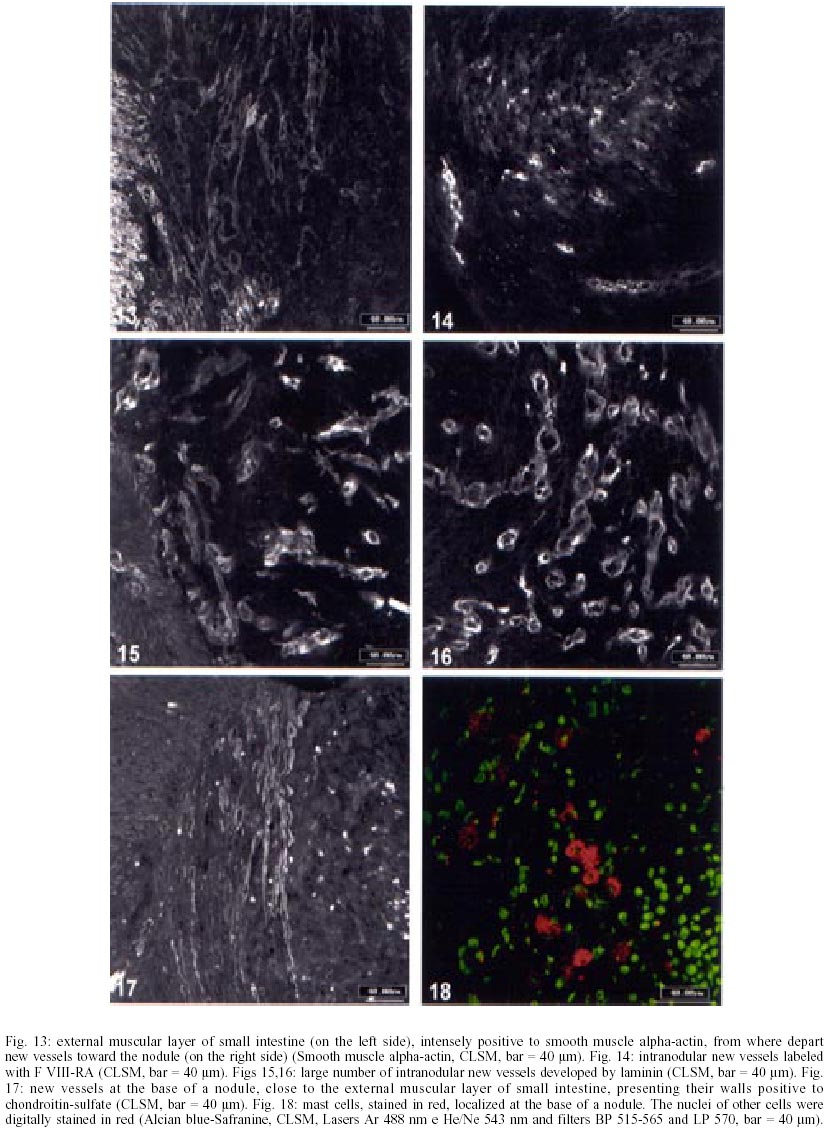
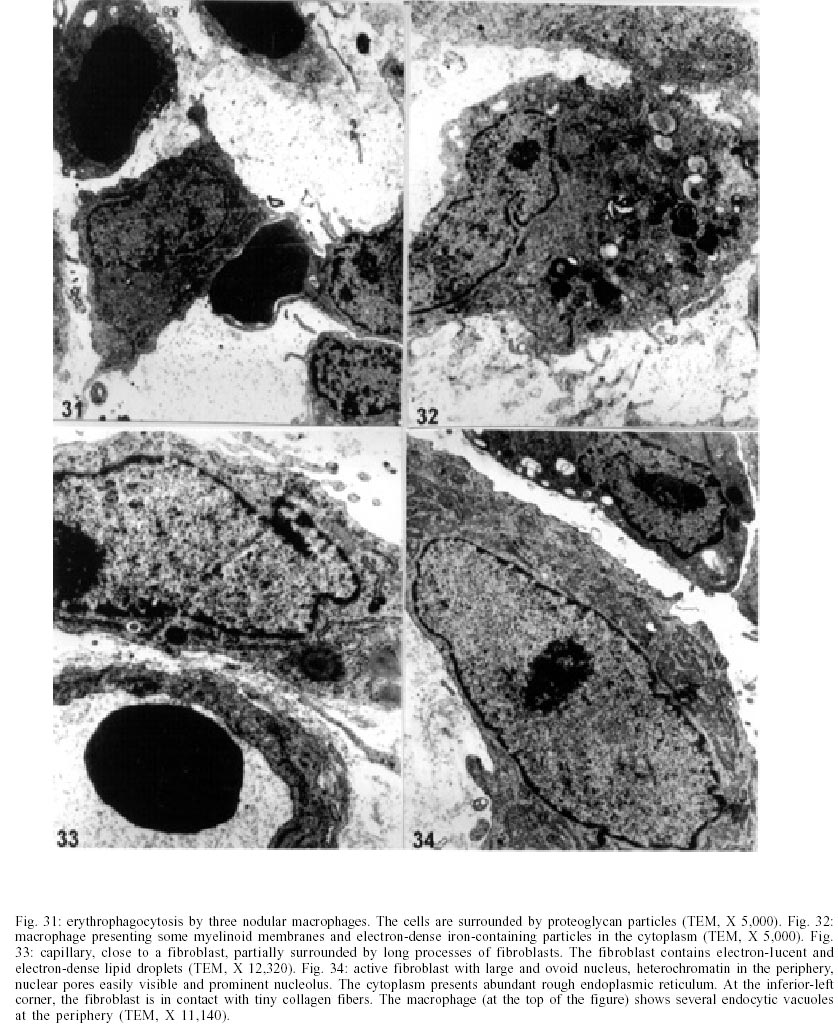
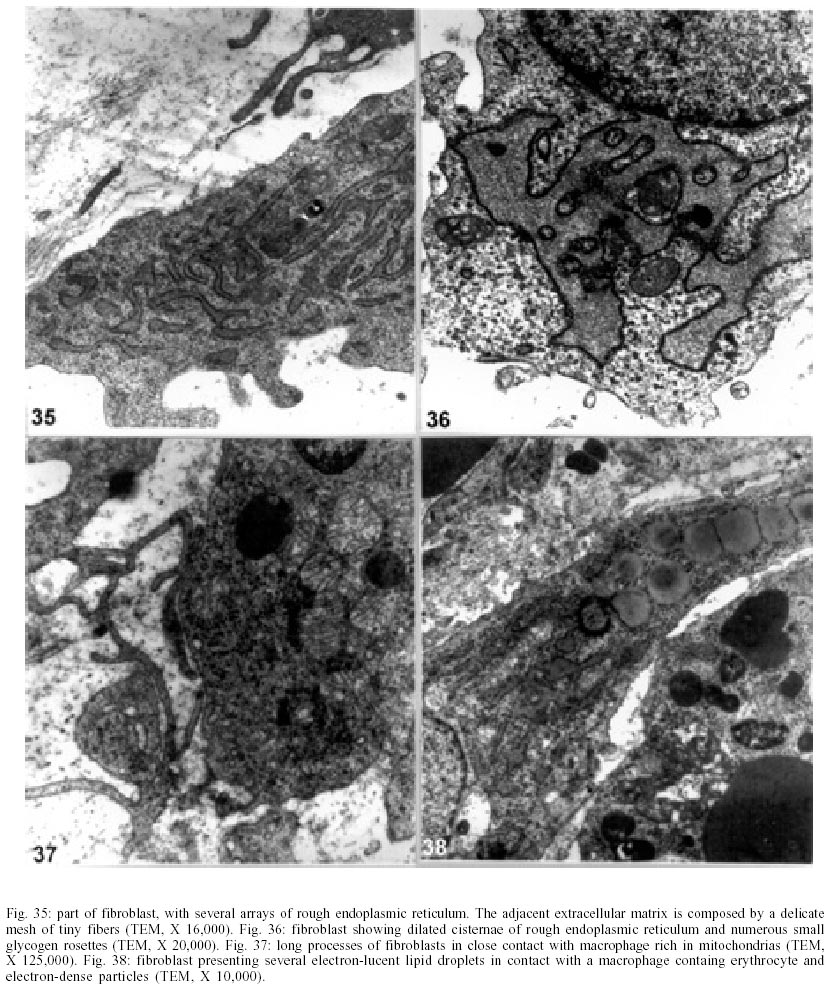
On days 55 up to 160 after infection, C. callosus developed prominent and diffuse intestinal subserosal nodules resulting in an overall granulated aspect on the intestinal surface (Figs 1, 2). The nodules were translucent with or without hemorrhage (Figs 1, 2) and by SEM showed a small pedicle and focal extravasation of red cells (Figs 3-6). When the nodules were isolated (Fig. 7) and cultivated in cell culture they radially originated from the third day onward, a mesh of fusiform cells (Figs 8, 9) that were very similar to the fibroblastic cells seen in paraffin section (Fig. 10). The nodules were composed by central granuloma(s) (Figs 25, 26) surrounded by intense and proliferative fibrovascular tissue (Figs 25-28). The fusiform cells presented prominent and ovoid nucleus, with small nucleolus (Fig. 10), and exhibited vimentin-positive (Figs 23, 24), desmin and smooth muscle alpha-actin-negative cytoskeleton. Strong deposits of carboxylated proteoglycans at the periphery of the cells, yielded a "tram track" appearance (Fig. 11). The vascular component of the nodules derived from proliferating vessels from the external muscular layer, which were smooth muscle alpha-actin (Fig. 13), Factor VIII-RA (Fig. 14), laminin (Figs 15, 16) and chondroitin-sulfate (Fig. 17) positive. Close to the vessels there were aggregates of mast cells, mainly in the base of the nodules (Fig. 18). The stroma of the nodules was diffusely rich in fibronectin (Fig. 19), collagen VI (Fig. 21), collagens I and III and in proteoglycans (carboxylated > sulfated). Tenascin and interstitial collagens were more intense under the serosal surface (Figs 20, 27, 28). The mesothelial cells were highly cytokeratin-positive and were limited to the serosa without migration toward the interior of the nodules (Fig. 22). The three-dimensional aspect of the interstitial collagens in the stroma of the nodules is shown in the Figs 25-30. Intermixed with the fibroblasts and vessels there were variable number of eosinophils, and large number of macrophages, sometimes presenting erythrophagocytosis, and containing ferric pigment (Fig. 12). The ultrastructural study confirmed the presence of macrophages with and/or without erythrophagocytosis (Figs 31, 32), vessels (Fig. 33) and showed active fibroblasts. These presented large and ovoid nucleus with peripheric heterochromatin and prominent nucleolus (Fig. 34); the cytoplasm exhibited abundant and sometimes dilated rough endoplasm reticulum (Figs 34-36). The cells were surrounded by extracellular elements (Figs 31, 32, 34, 35) and contacts between fibroblasts and macrophages were often seen (Figs 32, 37, 38). Some fibroblasts showed lipid vacuoles in the cytoplasm (Figs 33, 38). DISCUSSION
The intestinal nodules represent an exaggerated fibrovascular proliferation around periovular granulomas of S. mansoni. They are found at the interface between external muscular layer and intestinal serosa. The mechanisms of the angiogenesis in the nodules are still unknown. Smith (1961) and Schoefl (1963) claimed that mast cells and histamine are involved in capillary growth during injury. Indeed, we detected, mainly at the root of the nodules, a large number of connective tissue mast cells (CTMC) with safranin-positive granules, reflecting the presence of a highly N-sulfated polysaccharide, likely to be mature heparin (Fig. 18). Moreover, mast cells also release other substances [Adenosine diphosphate (ADP), 5-hydroxytryptamine (5-HT) and neutral proteases], and heparin has been repeatedly shown to have an important role in different steps involved in angiogenesis (Hudlická & Tyler 1986). Azizkhan et al. (1980) showed that heparin released from mast cell stimulates migration of capillary endothelial cells rather than their proliferation. Since migration of endothelial cells is an important factor in angiogenesis, mast cells may play an indirect role in the process, particularly in chronic inflammation. Heparin can also activate proteolytic enzyme degrading the extracellular matrix and can inhibit the proliferation of pericytes (Folkman et al. 1983). Usually pericytes restrain endothelial cell proliferation and differentiation (Orlidge & D'Amore 1987, Shepro & Morel 1993). Many cells present in the nodules are capable of producing angiogenic factors when their environment becomes hypoxic or inflammatory, including fibroblasts, monocytes/macrophages (Jackson et al. 1997), and maybe mesothelial cells. In fact, almost every growth factor and cytokine known to regulate angiogenesis can be produced by macrophages (Sunderkotter et al. 1994). Villaschi and Nicosia (1994) showed that fibroblasts could stabilize microvessel sprouts and stimulate angiogenesis from aortic explant cultures, a system that builds on co-culture. Mesothelial cells, in their turn, could produce many inflammatory mediators that have both direct and indirect angiogenic activities, such as: interleukins 1, 6 and 8 (IL-1, IL-6, IL-8), transforming growth factor beta (TGF-b), fibroblast growth factor (FGF) and granulocyte macrophage-colony stimulating factor (GM-CSF) (Zeillemaker et al. 1995). Goldsmith (1996) also detected angiogenic activity provoked by omental tissue, and Zhang et al. (1997) showed that vascular endothelial growth factor (VEGF) is the major angiogenic factor in omentum. The mesothelial cells could also up-regulate the fibrogenesis in the nodules, not only by the release of cytokines and growth factors, but also by the secretion of high levels of a plasminogen activator inhibitor PAI-1 (Rheinwald 1989). Human mesothelial cells in culture secrete high levels of this protein (Mr 45 KDa), which is also produced at equivalent levels by endothelial cells and kidney epithelial cells in culture, but at substantially lower levels by fibroblasts, and at almost undetectable levels by many other epithelial cell types (Rheinwald 1989). Plasminogen activator (PA) inhibitor (PAI-1) is an inhibitor of the serpin class of PA and plasmin, together with PAI-2 (60 KDa), protease nexin I (PN-1, 45 KDa), a2-antiplasmin, and aprotinin (7 KDa) (Saksela & Rifkin 1988, Laiho & Keski-Oja 1989). Tissue-type PA (TPA, 70 KDa) is involved in regulating the clotting process, and urokinase-type PA (UPA, 50 KDa) is active in fibrinolysis, as well as in biological remodeling, invasive processes, and can cleave fibronectin and plasminogen (Alexander & Werb 1991). Then the PAI-1 secretion can promote the accumulation of some stromal components in the nodules, by interference on extracellular matrix degradation. Parasite-derived products can also take part in the angiogenesis phenomenon. In fact, S. mansoni soluble egg antigen (SEA), in the absence of inflammatory cells, can stimulate proliferation of human umbilical vein endothelial cells (HUVE) in vitro (Freedman & Ottesen 1988). This effect appears to be due to up-regulation of human VEGF in endothelial cells by products secreted by schistosome eggs (Loeffler et al. 2002). The mesenchymal cells involved in the nodular fibrosis were characterized as fibroblasts and not as myofibroblasts due to the following aspects: they were negative for both alpha-smooth muscle actin and desmin and did not present by ultrastructural analysis folded or invaginated nucleus; abundant micropinocytotic vesicles; focal densities in the cytoplasm and a thin but usually distinct external lamina (Ghadially 1997). There was inhibition or desintegration of the external muscular intestinal layer, where the nodules were located, without evidence of subsequent modification of the smooth muscle cells to myofibroblast phenotype. The large amount of carboxylated and sulfated glycosaminoglycans observed in the nodules could be an inhibiting factor on proliferation of smooth-muscle cells. Heparin and heparan sulfate glycosaminoglycans are potent inhibitors of vascular smooth-muscle cell proliferation in vivo and in vitro (Reilly 1993). Data from the literature indicate that heparin has no effect on platelet derived growth factor (PDGF) binding, but markedly inhibits the mitogenic response of smooth-muscle cells to this same factor and to epidermal growth factor (EGF) (Reilly 1993). El-Awady et al. (2001) showed that S. hematobium SEA induced proliferation of human urothelial and bovine endothelial cells, but no effect was observed on smooth muscle cell lines. Similar prominent intestinal fibrovascular nodules were also seen by Rodrigues-Silva (1989) in Nectomys squamipes, natural and experimentally infected with S. mansoni. Portillo and Damian (1986) reported the presence of subserosal intestinal granulomatous nodules in a small New World monkey, the saddle-back tamarin Saguinus fuscicollis, subcutaneously infected with 150 S. mansoni cercariae, which were necropsied at 18 months. According to these authors, the nodules may represent a defensive response to the thin-walled intestine of the tamarin monkeys to the presence of a large number of sequestered eggs. Nodules that contained eggs were also found free in the peritoneal cavity, where they had become necrotic. Such apparent budding off of heavily egg-infiltrated tissues may represent an even more pronounced type of sequestration (Portillo & Damian 1986). Subserosal nodules resulting from confluent granulomas have been reported in humans and experimental animals with chronic S. mansoni infections (Kean & Dennis 1971, Marcial-Rojas 1971). It is important to point out that the fibrovascular nodules in C. callosus are different and larger than the granulomatous nodules observed by Domingo and Warren (1969) in Swiss albino female mice, studied after infection of S. mansoni for a period of 20 weeks and are also not directly related to local deposits of large number of eggs. On the contrary, in C. callosus model, there is a striking dissociation between the number of eggs per nodule and the pronounced perigranulomatous fibrovascular reaction. In this sense, pathologists have also been intrigued by large fibrous tumors caused by S. japonicum infection (bilharziomata) containing relatively few eggs (Lichtenberg et al. 1971). In conclusion, the intestinal nodules in C. callosus are fibrovascular structures stimulated by local granulomas close to the serosa. They constitute an excellent and accessible model to study fibrogenesis and angiogenesis dependent on S. mansoni eggs. The fibrogenic activity is fibroblastic and not myofibroblastic-dependent. The angiogenesis is so prominent that causes haemorrhagic ascites. At least, in this model, the hypervascularity does not reflect inflammatory vasodilatation of preexisting host vessels, but is the outcome of angiogenesis. This model allows investigating the following questions: (1) What are the local factors that simultaneously stimulate fibrous and vascular proliferation? Both events occur concomitantly in the human schistosomiasis, validating the importance of this experimental model; (2) What is the participation of mesothelial cells in this type of intestinal fibrosis and angiogenesis? (3) What are the local epigenetic factors involved in the angiogenesis-granuloma dependent, i.e., why this angiogenesis is not universally expressed in all the organs with schistosomal granulomas? (4) Do the inhibitors of angiogenesis interfere on fibrogenesis process? REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02200f7-12.jpg] [oc02200f13-18.jpg] [oc02200f1-6.jpg] [oc02200f31-34.jpg] [oc02200f35-38.jpg] [oc02200f25-30.jpg] [oc02200f19-24.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}