 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(Suppl. I) October 2002, pp. 129-142 Natural Schistosoma mansoni Infection in Nectomys squamipes: Histopathological and Morphometric Analysis in Comparison to Experimentally Infected N. squamipes and C3H/He mice Michele Costa-Silva/++, Rosângela Rodrigues-Silva*, Maarten Hulstijn, Renata Heisler Neves, Mônica de Souza Panasco**/++++, Henrique Leonel Lenzi**/+++, José Roberto Machado-Silva/+/+++
Departamento de Patologia e Laboratórios,
Faculdade de Ciências Médicas, Universidade do Estado do Rio de
Janeiro, Rua Prof. Manuel de Abreu 444, 5o andar, 20550-170 Rio de
Janeiro, RJ, Brasil *Laboratório de Helmintos Parasitos de Vertebrados,
Departamento de Helmintologia **Departamento de Patologia, Instituto Oswaldo
Cruz-Fiocruz, Rio de Janeiro, RJ, Brasil This work is part of a MSc Thesis in Morphology, Instituto de Biologia, Uerj, RJ, Brasil. Received 18 June 2002 Code Number: oc02201
Histopathologic and morphometric (area, perimeter, major and minor diameters) analysis of hepatic granulomas isolated from twelve naturally infected Nectomys squamipes were compared to four experimentally infected ones and six C3H/He mice. Liver paraffin sections were stained for cells and extracellular matrix. Both groups of N. squamipes presented peculiar granulomas consisting predominantly of large macrophages, full of schistosome pigment, characterizing an exudative-macrophage granuloma type, smaller than the equivalent granuloma type in mouse. Naturally infected animals exhibited granulomas in different stages of development, including large number of involutional types. Morphometric analysis showed that all measurements were smaller in naturally infected animals than in other groups. The results demonstrated that both N. squamipes groups reproduced, with small variations, the hepatic granuloma aspects already described in cricetidium (Calomys callosus), showing a genetic tendency to set up strong macrophage responses and small granulomas. Unexpectedly, natural infection did not engender distinguished histopathological characteristics distinct from those derived from experimental single infection, showing changes predominantly secondary to the duration of infection. It appears that the variability of the inocula (and the number of infections?) interfere more with the quantity than with the quality of the pathological changes, denoting some morpho-functional determinism in the response to schistosomal infection dependent on the animal species.
Keys words: Schistosoma mansoni - Nectomys squamipes - C3H/He mice - hepatic granuloma - histopathology - morphometry Several authors have shown that rodents in Africa or in Neotropical areas can harbor Schistosoma mansoni infection (Théron et al. 1992, Alarcón de Noya et al. 1997, Morand et al. 1999, Duplantier & Sène 2000). In Brazil, several reports indicated that Nectomys squamipes Brants 1827 (Rodentia: Sigmondontinae) has an important role in the transmission of this helminthiasis depending on special ecological conditions (Rey 1993, D'Andrea et al. 2000). N. squamipes, semiaquatic species of Sigmodontinae, is distributed along watercourse banks and has a wide geographic range coincident with S. mansoni endemic areas (Ribeiro et al. 1998). Laboratory studies have demonstrated that N. squamipes is highly susceptible to experimental S. mansoni infection (Rodrigues-Silva 1988, Souza et al. 1992, Maldonado Jr. et al. 1994, Ribeiro et al. 1998). It presents a well-balanced relationship with the parasite, since life-long infections do not affect its life span (Rodrigues-Silva et al. 1992) or its reproductive capacity (D'Andrea et al. 2000), therefore being considered a permissive host (Machado-Silva et al. 1997). Mature eggs of S. mansoni retained in the intestine and liver tissues of immunological competent hosts induce granuloma formation, which regulation and morphological aspects vary among different mice strains and animal species (Cheever et al. 1998). For instance, hepatic granulomas of Calomys callosus (Rodentia: Cricetidae) are 27% smaller than granulomas from albino mice at the same time of infection (Lenzi et al. 1995). The granuloma variability is due to the fact that this organized structure is composed by host and parasite components, functioning as a hybrid interface between two different phylogenetic beings, depending, therefore, on the host epigenetic characteristics (Lenzi et al. 1998a). Most studies of the granulomatous reaction seen in literature, except in human cases, make use of experimentally infected animals, introducing into the analysis inexorable artificial variables. Here we present an interesting animal model (N. squamipes) which takes part in the maintenance of S. mansoni life-cycle in one endemic area, pointing out the qualitative and quantitative histopathological aspects of hepatic granulomas following natural infection. The data were compared with granulomas from N. squamipes and C3H/He mice born and infected under laboratory conditions. MATERIALS AND METHODS
Animals and S. mansoni infection - Twelve specimens of naturally infected N. squamipes (2n = 56 chromossomes) (Bonvicino et al. 1996) were captured in the Pamparrão Valley, District of Sumidouro, State of Rio de Janeiro, Brazil, and eleven were killed in accordance with animal ethical practice seven days after their capture (Rodrigues-Silva et al. 1992). One animal, after the capture, was maintained in the laboratory for 490 days before its necropsy. From the feces of these animals, a S. mansoni isolate was obtained (R isolate), and the methodology applied for its isolation and maintenance in laboratory was already presented in another publication (Machado-Silva et al. 1994). Briefly, specimens from a sympatric Biomphalaria glabrata colony, kept under laboratory conditions, were exposed to ten miracidia. The snails were maintained in glass aquariums and fed ad libitum with fresh lettuce (Lacttuca sativa). The cercariae eliminated after exposure of snails to a light source were used for experimental infection. Four specimens of N. squamipes and six C3H/He mice were infected by percutaneous exposure to 500 and 50 cercariae respectively, and were killed in the same manner as naturally infected ones at 52 and 62 days after infection, respectively. Histopathological analysis - Liver fragments taken from the right lobule of each animal, was fixed in Carson's modified Millonig's phosphate-buffered formalin, pH 7.4 (Carson's formalin-Millonig) (Carson et al. 1973) and embedded in paraffin. Sections (5 µm) were stained with hematoxilin and eosin (H&E); PAS-Alcian Blue, pH = 1.0 and 2.5; Lennert's Giemsa (Lennert 1978); Gomori's reticulin stains, Sirius red pH = 10.2 (eosinophils) (Bogomoletz 1980, Luque & Montes 1989, Vale et al. 1997); Masson's trichrome, Alcian blue/safranin (mast cell subpopulation) (Strobel et al. 1981) and Picrosirius ± polarization microscopy (Junqueira et al. 1979) and Weigert resorcin-fuchsin ± oxidation with potassium peroxymonosulfate (OxoneÒ monopersulfate compound, Sigma 22,803-6). Digital images were captured with Hamamatsu color chilled 3 CCD camera (model C-5810), stored with tagged-image file format (TIF) and printed in Codonics NP-1600 Photographic dye-sublimation Printer. Morphometric analysis - Brightfield microscopy (Olympus BX50) was used to evaluate the sections. Images were captured by an analogue camera (Sony, 640 x 480 pixels, RGB) and transferred to a computer containing software for image analysis (Image Pro Plus - Media Cybernetics). Morphometric analysis of the granulomas was performed measuring the following parameters: area, perimeter, major and minor diameter. Only granulomas with eggs visible in the centre were analyzed. Statistical analysis - One-way ANOVA was applied using the post hoc Tuckey test to compare data from each group individually. Measurements with p values £ 0.05, were considered significantly different (Vieira 1991). RESULTS
Histopathological results
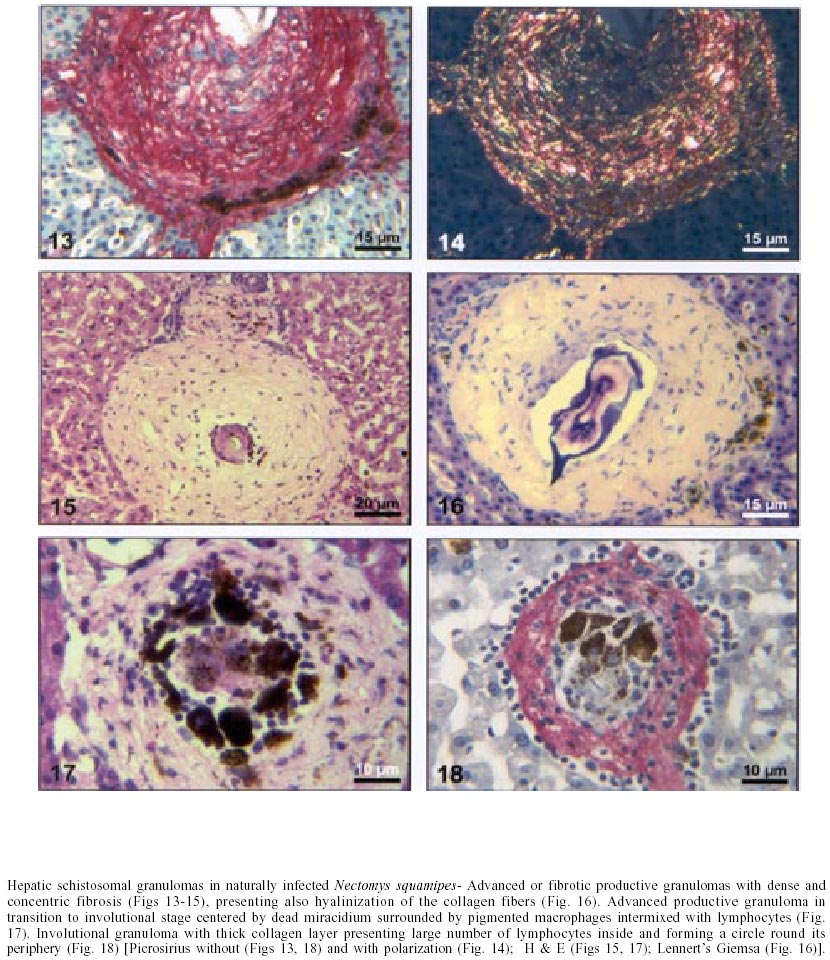
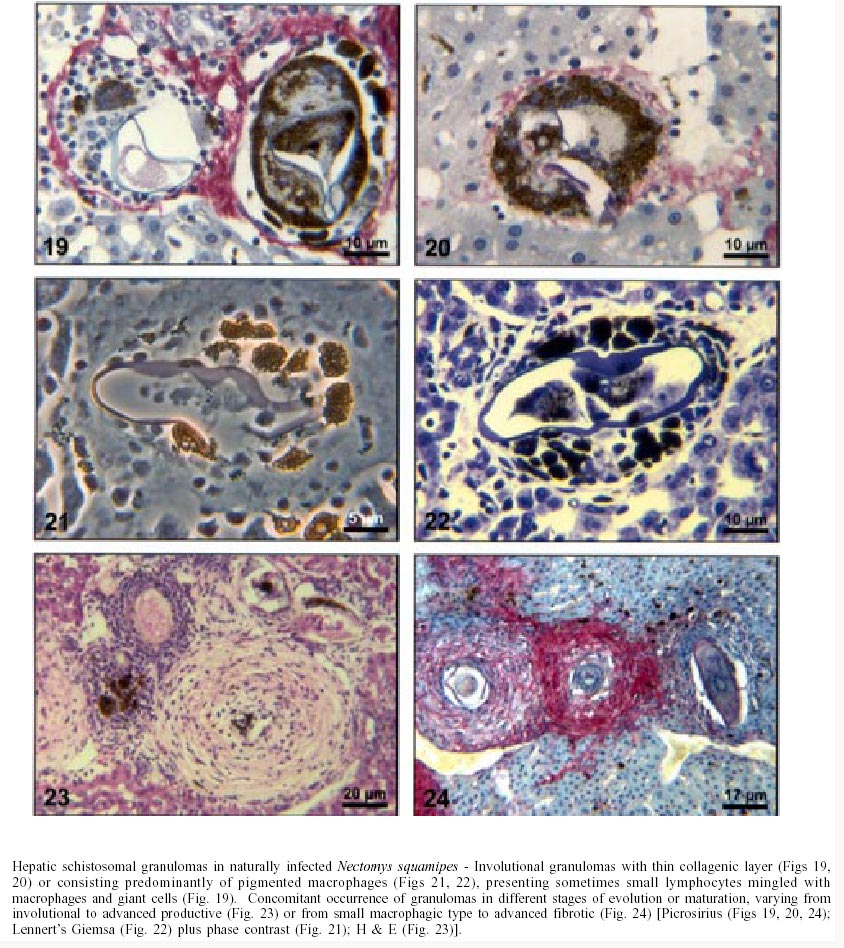
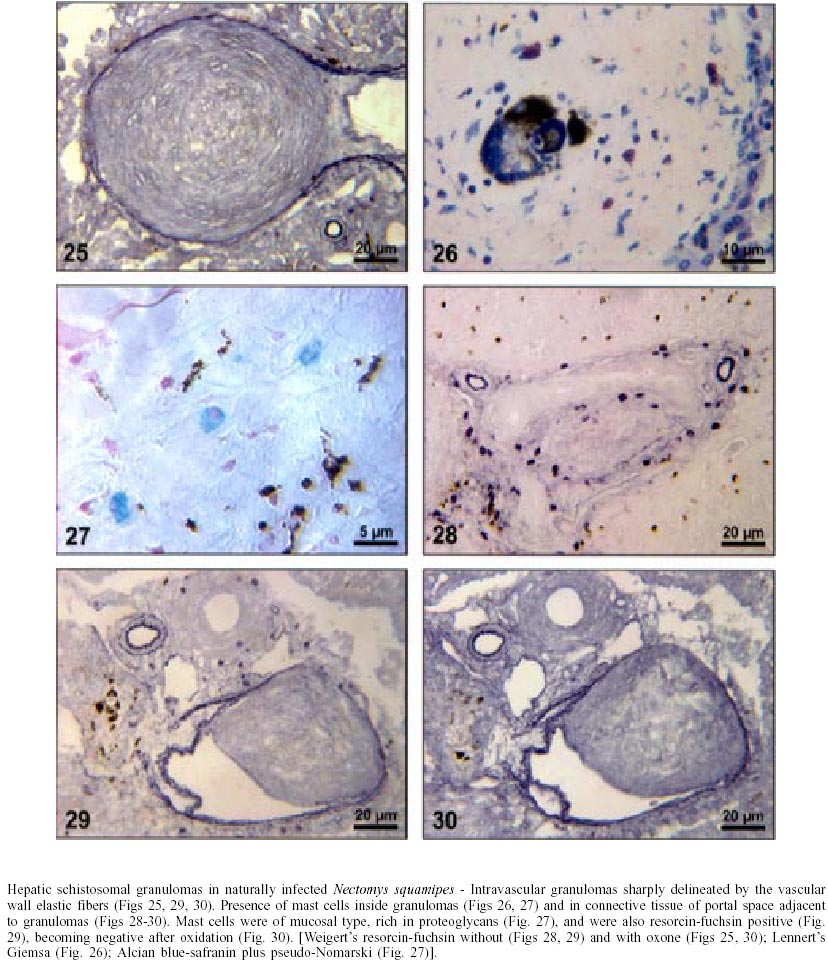
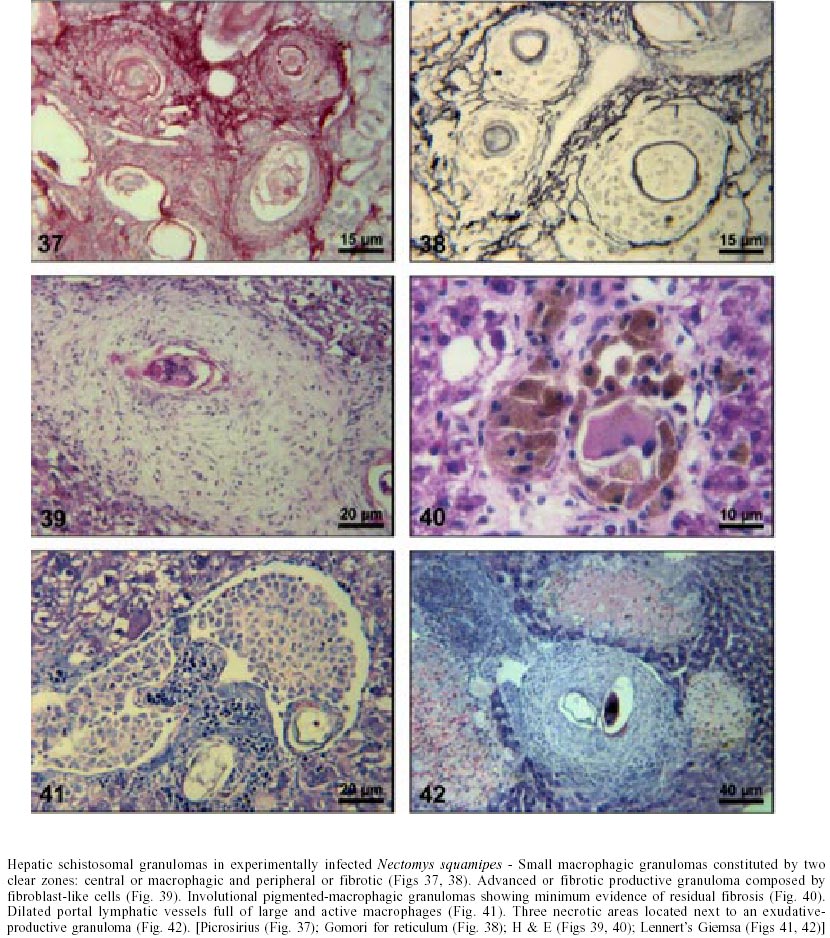
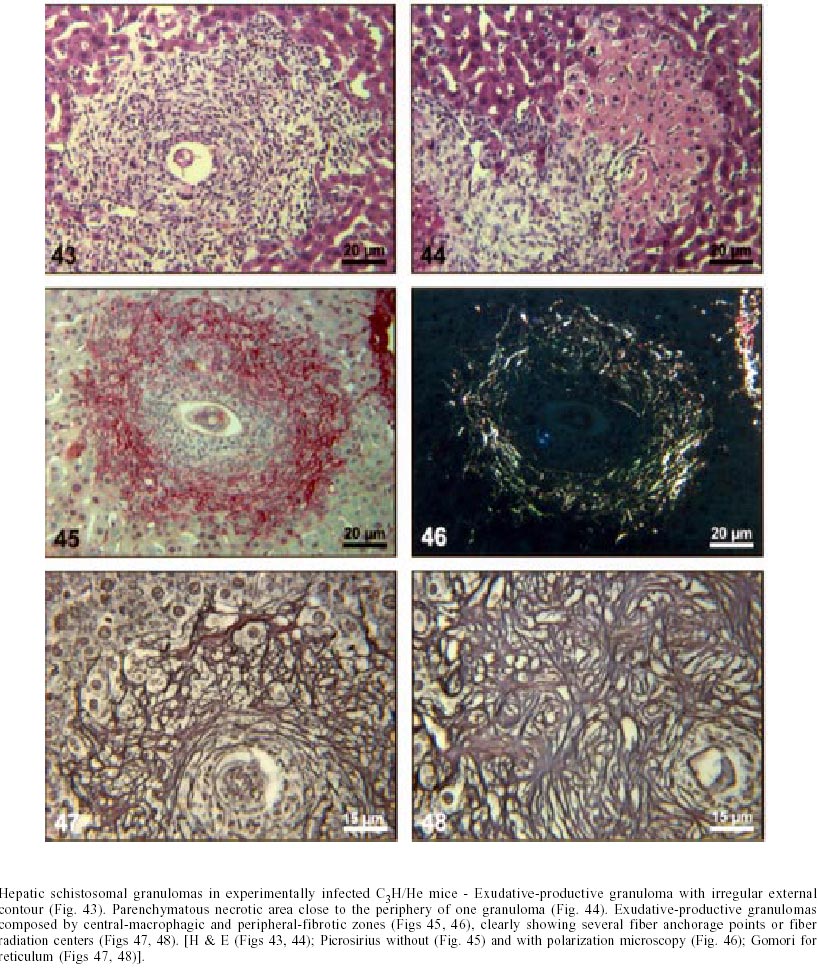
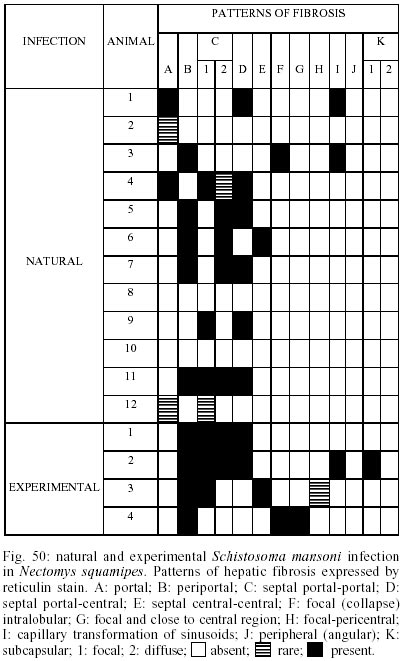
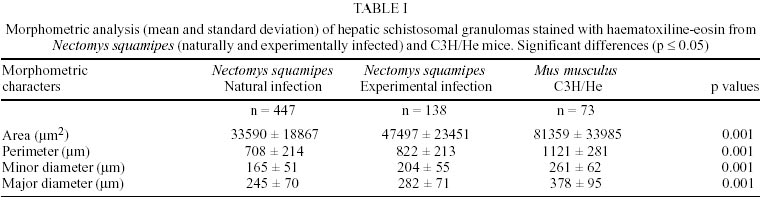
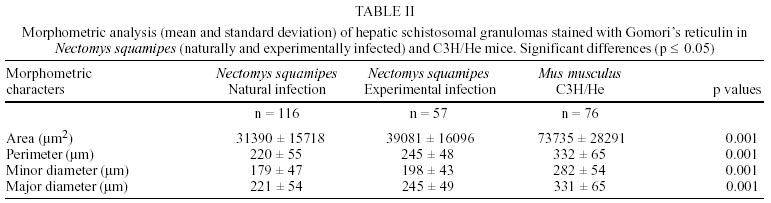
Naturally infected N. squamipes - Livers from naturally infected N. squamipes showed peri-ovular lesions in pre-granulomatous stages, characterized by exudative (Fig. 1), exudative-necrotic reactions (Fig. 2), and clear granulomatous stages in different developmental phases such as exudative-productive (Fig. 3), productive (Fig. 4) and involutional subtypes (Figs 17-22). The exudative-productive and involutional granulomas predominated in number and the former were composed of a large macrophagic central layer, surrounded by an external one consisting of lymphocytes, plasma cells and pigmented macrophages with variable number of eosinophils (Fig. 3). These two layers were sharply demarcated by a reticular scaffold, forming a mesh with concentricity of the fibers (Figs 5, 6). Sometimes, in large portal exudative-productive granulomas three layers were clearly defined by the arrangement of reticular fibers: inner, internal or paucifibrillar zone, which consisted of macrophages; the middle or paracentral zone rich in fibers and the external zone, predominantly cellular with scarce amount of fibers (Fig. 6). These portal granulomas expressed large amounts of interstitial collagens marked by Picrosirius in brightfield (Figs 7, 9, 11, 13) or in polarization microscopies (Figs 8, 10, 12, 14). The intragranulomatous collagenic disposition presented two patterns: one central or centrifugal (Figs 7-10) and an another concentric and peripheral, that tended to make a delimiting fibrotic pseudocapsule (Figs 7-10) which became exacerbated in productive granulomas, showing a centripetal tendency or a peripheral to central decrescent gradient (Figs 11-14). The advanced or fibrotic productive granulomas were spherical composed mainly by fibroblast-like cells (fibroblasts or myofibroblasts?), displaying sometimes hyalinization of collagen stroma (Figs 15, 16). Involutional stages of granulomas were frequent, varying from transitional forms between advanced-productive to early involutional (Fig. 17), involutional with thick (Fig. 18) or thin (Figs 19, 20) collagen layer or predominantly pigmented macrophagic type (Figs 21, 22). Small lymphocytes were often detected in these types of involutional granulomas, located together with the pigmented macrophages or making a peripheral halo (Figs 17-20, 22). The variegated granuloma types cohabited simultaneously in the liver of each individual animal (Figs 23, 24). Some granulomas developed intravascularly in portal veins, displacing the vascular wall to the periphery (Figs 25, 29, 30). The expression of high or low ph proteoglycans in the granulomas was not homogeneous, being more intense in the granulomas of animals with morphological evidence of more chronic infection. Numerous mast cells, sometimes in degranulation were detected in granulomas or mainly in the connective tissue of portal spaces close or not to granulomas (Figs 26-29). They were alcian blue positive in Strobel stain (Fig. 27), being also more alcian blue positive in pH 1.0 than in pH 2.5. They stained highly dark with resorcin-fuchsin without oxidation (Fig. 29), becoming weak or negative after oxidation with oxone (Fig. 30). Portal spaces, beyond mast cells, exhibited plasma cells and less frequently eosinophils, macrophages with or without schistosome pigment and lymphoid aggregates. Portal and periportal fibrosis, and bridging fibrous septa connecting portal spaces or portal spaces to central zones were changeful and usually the fibrosis was related to the presence of granulomas. Some portal spaces showed dilated lymphatic vessels. The Kupffer's cells were hypertrophic, containing variable amount of schistosome pigment, and small and isolated parenchymatous necrosis was seen close to granulomas. All granulomas types were seen in naturally infected animals although productive-fibrotic was predominant. Experimentally infected N. squamipes - The four experimentally infected animals with 52 days of infection showed a limited pattern of granulomas. The animals displayed a weak or initial peri-ovular reaction, exudative (Fig. 31) and exudative-necrotic pre-granulomatous stages, exudative-productive (Figs 32, 34), pure macrophagic (Fig. 33), small fibrotic-macrophagic (Figs 37, 38), and more scarcely productive (Fig. 39) and involutional (Fig. 40) granulomas. The macrophagic variations of granulomas were the predominant types. The granulomas expressed simultaneously high and low sulphated proteoglycans, and presented less amount of collagen than in the naturally infected animals (Figs 35, 36). The patterns of portal and septal fibrosis and the Kupffer's cells changes were similar to those described in the naturally infected group. However, two animals showed some portal lymphatic vessels with lumen prominently expanded and full of macrophages (Fig. 41), and, in general, the animal of this group presented more focal parenchymatous necrosis (Fig. 42). Experimentally infected C3H/He mice - Mice with 62 days of infection presented exudative peri-ovular lesions and a clear predominance of exudative-productive granulomas, without evidence of more advanced stages of the granulomatous reaction (Fig. 43). Like the N. squamipes, focal areas of parenchymatous necrosis arose close to some granulomas (Fig. 44). The pattern of architectural arrangement of collagen fibers inside the granulomas tended to define a trellis-like or storiform disposition, making radial extensions to adjacent parenchyma (Figs 45-47) and presenting several fiber anchorage points or fiber radiation centers (Fig. 48). Schematic comparative views between granuloma types and patterns of extragranulomatous fibrosis observed in natural and experimentally infected N. squamipes groups are displayed in Figs 49 and 50. Morphometric results - Morphometric analysis showed that all measurements of liver granulomas evaluated in sections stained with HE or Gomori's reticulin were smaller in naturally infected animals than in the other two groups. Mice displayed the largest values while measurements of experimentally infected N. squamipes were intermediate (Table I, II). Comparing all parameters among the three groups, there were highly significant differences between N. squamipes and the mice (p < 0.001). DISCUSSION
This paper focused on the morphological aspects of hepatic changes in naturally S. mansoni infected N. squamipes, showing evidence of active infection expressed by frequent viable miracidia inside the eggs and recent pre-granulomatous and granulomatous peri-ovular reactions. The general morphological aspects of the granulomas in naturally infected animals when compared with the experimentally infected ones indicated, based on the characteristics of the more advanced granulomatous stages, that the infection was more chronic and certainly surpassed 52 days of infection. However, the data did not provide any conclusion whether the captured animals were infected only once or many times under natural conditions. Experimental studies have shown that N. squamipes is susceptible to reinfection (Maldonado Jr. et al. 1994). Surprisingly, there were not meaningful differences between the granuloma aspects of naturally infected N. squamipes and chronically infected C. callosus, which were infected only one time with 70 S. mansoni cercariae (Lenzi 1998). In this work, N. squamipes with 52 days of infection exhibited more homogeneous granulomas due to the short time of infection, although the granulomas were different from the mice, being predominantly of exudative-macrophage type. The survival of one animal naturally infected for more than 400 days in laboratory conditions (it was killed at 490 days after capture) (Rodrigues-Silva 1988) emphasized the resistance of N. squamipes to the infection and its putative weightiness to keep on maintaining the S. mansoni cycle in endemic areas where they play role as an alternative vertebrate host (D'Andrea et al. 2000). N. squamipes like C. callosus (Lenzi et al. 1995), presented peculiar granulomas consisting predominantly of large macrophages, many of them full of schistosome pigment, characterizing a exudative-macrophage granuloma type, usually smaller than the equivalent granuloma type in mouse. They were composed by large macrophages with variable number of eosinophils, surrounded or not by mononuclear cells or were small and constituted by a macrophagic central layer surrounded by a thin collagenous and reticular pseudocapsule (Figs 3, 33, 37, 38). The frequent presence of intravascular granulomas as was seen in C. callosus (Lenzi et al. 1995) may be due to a peri-ovular reaction inside the vessels with less granulocytes than in mice, poor in proteolytic enzymes responsible for corrosion of the vascular walls. The presence of small parenchymatous necrotic foci in the presence of very well configured granulomas in both experimentally infected groups (N. squamipes and C3H/He mice) suggested that this event that happens more in acute phase of infection is not dependent on the granuloma efficiency to circumvent egg antigens and its pathogenesis is still unknown. Large macrophages full of Schistosoma pigment in the granulomas, mainly of the involutional types, characterized another feature of N. squamipes infection that was similar to C. callosus (Lenzi et al. 1995, Lenzi 1998). Recently, this pigment was unequivocally identified as haemozoin (Hz), and similar to Plasmodium and Rhodnius prolixus, S. mansoni produces Hz in order to detoxify the haem derived from haemoglobin digestion (Oliveira et al. 2000a). Therefore, Hz formation in the blood-feeding organisms seems to be an important mechanism to prevent generation of free radicals by haem released upon haemoglobin digestion (Slater et al. 1991, Oliveira et al. 1999, 2000a,b, Chen et al. 2001). Oliveira et al. (2002) demonstrated that the pro-oxidant activity of Hz depended on the crystal's size. Capron et al. (1965) proposed a causal relationship between pigment deposits and fibrogenesis in liver. However, Andrade and Andrade (1965), Stenger et al. (1967), Grimaud et al. (1976) and Lenzi et al. (1995, 1998b) have indicated absence or low fibrogeneis around the pigmented-loaded cells. In fact, Hz effects on the granuloma development and regulation were not well explored in the S. mansoni pathogenesis, since Hz has been involved in many biological activities such as impairment of phagocytic function (Schwarzer & Arese 1996), induction of lipoperoxidation (Schwarzer et al. 1996, Arese & Schwarzer 1997), 4-hydroxynonenal (HNE) and hydroxyeicosatetraenoic acids (HETEs) production (Green et al. 1996), immunomodulations of macrophages affecting antigen processing, chemokine and cytokine secretion (Pichyangkul et al.1994, Green et al. 1996, Schwarzer et al. 1998, Scorza et al. 1999). The frequent presence of multinucleated giant cells engulfing egg detritus in involutional N. squamipes granulomas was considered as indicative of their effect as schistosome egg scavenger after miracidia death instead of destructive cells. Multinucleated giant cell formation was also observed in olive baboon (Papio cynocephalus anubis) hepatic granulomas and, differently from our point of view, was presented as a cellular arrangement particularly efficient in bringing about the destruction of schistosome eggs and subsequent resolution of the egg granuloma without fibrosis (Farah et al. 2000). N. squamipes involutional granulomas presented peculiar distribution of lymphocytes that were mixed with or around pigmented macrophages (Figs 17, 18, 19, 20) and/or formed a ring around the granulomas (Fig. 18). So far there is no lymphocyte marker to Cricetidae family of animals making it impossible to define the lymphocyte sub-populations. However, the lymphocytes in involutional granulomas can act as macrophage activators inducing the expression of tumor necrosis factor-alfa and matrix metalloproteinases (Fayyazi et al. 2000), accelerating the granuloma degradation and absorption. Unlike C. callosus (Lenzi et al. 1995), another member of Cricetinae (Sigmodontinae) subfamily, N. squamipes presented a large number of mucosal mast cells in portal spaces and inside granulomas mainly of the proliferative fibrotic type (Figs 26-29). Fibroblasts are believed to promote mast cell survival and differentiation through the production of membrane-bound and soluble stem cell factor (SCF), which interact with c-kit on the surface of mast cells (Levi-Schaffer et al. 1986, Vliagoftis et al. 1999); they can also interact with mast cells through connexins, since mast cells express CX 43 an CX 32 (Oliani et al. 1995). Otherwise, mast cell-derived mediators and mast cell-related neutral proteases, tryptase and carboxypeptidase are potent fibrogenic factors that are capable of inducing fibroblast proliferation and type I collagen synthesis (Gailit et al. 2001). Mast cell mediators can also play a role in the regulation of myofibroblasts differentiation and function (Horobin & Flemming 1980). Mast cells exhibited staining characteristics similar to elastic fibers, showing attenuation and even null reaction after oxidation by oxone probably due to conversion of disulfide bridges to anionic sulphonic acid derivatives, changing the affinities to the basic dye compounds of the Weigert's method (Figs 29, 30) (Bogomoletz 1980, Avraham et al. 1989). The extragranulomatous and/or portal fibrosis in N. squamipes liver, like in mice, never acquired Symmer fibrosis pattern, but sometimes produced local obstruction of portal lymphatic vessels causing focal lymphangiectasis full of macrophages (Fig. 41). The lymphatic vessels in portal tracts drain the Disse spaces and although the lymph flows in the same direction as the bile, opposite to that of the blood (Thung & Gerber 1992), there was no evidence of biliary obstruction. In conclusion, the results showed that N. squamipes, natural and experimentally infected with S. mansoni, reproduced, with small variations, the hepatic granuloma aspects already described in cricetidium (C. callosus) (Lenzi et al. 1995, Lenzi 1998b), showing a genetic tendency to set up strong macrophage responses and small granulomas. Unexpectedly, natural infection did not engender distinguished histopathological characteristics distinct from those derived from experimental single infection, showing changes predominantly secondary to the duration of infection. It appears that the variability of the inocula (and the number of infections?) interfere more with the quantity than with the quality of the pathological changes, denoting some morpho-functional determinism in the response to schistosomal infection dependent on the animal species.
ACKNOWLEDGEMENTS
To Ms Luzia de Fátima Gonçalves Caputo from Pathology Department, Oswaldo Cruz Institute, for her expert technical assistance. To Genilton José Oliveira and Heloísa Maria Nogueira Diniz from Education Department, Oswaldo Cruz Institute, for preparation of figures. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02201f50.jpg] [oc02201f37-42.jpg] [oc02201f1-6.jpg] [oc02201t1.jpg] [oc02201f49.jpg] [oc02201t2.jpg] [oc02201f13-18.jpg] [oc02201f43-48.jpg] [oc02201f7-12.jpg] [oc02201f19-24.jpg] [oc02201f31-36.jpg] [oc02201f25-30.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}