
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 7, 2002, pp. 917-940
|
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(7), October
2002, pp. 917-940
Experimental Models
of Schistosoma mansoni Infection
Allen W Cheever, Jane A Lenzi*, Henrique
L Lenzi*, Zilton A Andrade**/+
Biomedical Research Institute, MD
20852-1709, USA *Departamento de Patologia, Instituto Oswaldo Cruz-Fiocruz,
Rio de Janeiro, RJ, Brasil **Laboratório de Patologia Experimental, Centro
de Pesquisas Gonçalo Moniz-Fiocruz, Rua Valdemar Falcão 121, 40295-001
Salvador, BA, Brasil
+Corresponding author. Fax:
+55-71-356-4320, Ext. 206.
E-mail: zilton@cpqgm.fiocruz.br
Received 15 August 2002
Accepted 18 September 2002
Code Number: oc02214
Experimental models of Schistosoma
mansoni infections in mammals have contributed greatly to our understanding
of the pathology and pathogenesis of infection. We consider here hepatic and
extrahepatic disease in models of acute and chronic infection. Experimental
schistosome infections have also contributed more broadly to our understanding
of granulomatous inflammation and our understanding of Th1 versus Th2 related
inflammation and particularly to Th2-mediated fibrosis of the liver.
Key words: Schistosoma mansoni
- experimental models - schistosomiasis
Experimental schistosome infections
of laboratory animals have frequently been used to model the anatomo-pathologic
and pathophysiologic features of the infection in humans as well as for the
study of immunity and treatment. We concentrate here on the anatomic and parasitologic
features of various models and on the use of models to address mechanisms of
pathogenesis. Reviews dealing with immunopathology (Lukacs & Boros 1993,
Wynn & Cheever 1995, Cheever & Yap 1997, Fallon 2000) and with immunization
and resistance to reinfection (James 1995, Richter et al. 1995, Coulson 1997,
Waine & McManus 1997, Bergquist & Colley 1998) have been recently published.
Schistosoma mansoni matures
over a 5 week (wk) period in permissive hosts such as the mouse and egg laying
begins at that time. Most pathology in schistosome infected animals is attributed
to the host's reaction to the eggs which is maximal by the 8th wk of infection.
Granulomas are composed principally of macrophages, eosinophils and lymphocytes
with the proportion of cells varying in different organs (Weinstock & Boros
1983a). Natural killer cells may comprise over 20% of cells in the granuloma
(Remick et al. 1988), but these produced little IFN-g
(Rakasz et al. 1998). Mast cells are infrequent in 8-wk granulomas in most mouse
strains and become more frequent in chronic infections (Weinstock & Boros
1983b) and these may be important because they secrete fibrogenic mediators
and interact with hepatic stellate (Ito) cells (Brito & Borojevic 1997).
Chesney et al. (1998) described the infiltration of circulating "fibrocytes"
into granulomas and speculate that these cells may be important for attracting
CD4+ lymphocytes as well as for collagen formation.
After the 8th wk of infection there
is downmodulation of the immune reaction and granulomas around recently deposited
eggs become progressively smaller (Andrade & Warren 1964, Chensue &
Boros 1979). Although the response to new eggs is downregulated, cumulative
damage occurs as older lesions involute to leave fibrous scars. Thus the rate
of damage decreases but accumulated damage may increase, the balance being determined
by the variable ability of the host to kill worms, to inhibit worm fecundity
and to destroy eggs and repair tissue damage. The rhesus monkey does all these
things very well and shows no residual damage after the infection has cleared
or been treated (Cheever & Powers 1969, 1971). The baboon and cercopithecus
monkey destroy eggs rapidly and repair tissue damage (or perhaps never synthesize
much collagen) but kill worms slowly and inhibit oviposition slightly (Cheever
& Duvall 1974) and there is little cumulative damage. In the chimpanzee
worm fecundity is maintained and hepatic collagen and obstructive portal lesions
accumulate (Sadun et al. 1970). Major findings in the varied species used to
examine S. mansoni infections are summarized in Tables I,
II, III,
IV.
Schistosome infected animals are
exposed to antigens from the developing worms during the 5 wk before egg deposition
begins. The interpretation of immune reactions to the eggs is complicated by
this previous exposure to antigens, including antigens cross-reactive with egg
antigens (Lukacs & Boros 1991). Unisexually infected mice and mice sensitized
to many worm antigens are also sensitized to egg antigens and have an augmented
and accelerated response to injected eggs (Cheever et al. 1997, Jacobs et al.
1997a, 1998c), in unexplained contrast to the report of Warren and Domingo (1970).
The intravenous injection of eggs
initiates synchronous granulomas in the lung of a host which may be naive to
schistosome antigens or treated in a defined fashion. The subsequent development
of the granulomas is not entirely synchronous, but these lesions are more easily
studied than the completely non-synchronized granulomas resulting from infection.
The lung model is not, however, a substitute for the study of infected animals.
The antigenic quality of the eggs injected may affect both the size of the granuloma
and the effects of treatment on the granuloma (Eltoum et al. 1995).
Beads coated with schistosome egg
antigens, antigen fractions or recombinant antigens may also be injected intravenously
or used in vitro (Parra et al. 1991, Oliveira et al. 2000). Injection of beads
or eggs into the portal vein has been employed less frequently. Some investigators
have found naive mice nonresponsive to eggs injected into the portal vein (Leptak
& McKerrow 1997) and that portal (Cuison et al. 1995) or enteric (Weinstock
et al. 1985) injection of eggs induced tolerance to eggs subsequently injected.
Others have not noted this effect and have used portal injection as they would
the lung model (Edungbola & Schiller 1979, Raso et al. 1983, Eltoum et al.
1995, Jacobs et al. 1997a, 1999).
Granulomas in the mouse lung induce
much less fibrosis than granulomas in the liver and although it is possible
to study fibrosis in the lung model (Boros et al. 1983, Metzger & Peterson
1988), the high background levels of matrix and the low levels of fibrosis induced
(Cheever et al. unpublished) complicate this use of the lung model. Examination
of collagen mRNA levels in the lung (Warmington et al. 1999) will doubtless
be increasingly used as will determination of mRNA for proteases.
GENERAL CONSIDERATIONS IN THE
INTERPRETATION OF MODELS OF SCHISTOSOME INFECTIONS
A. The intensity of experimental
schistosome infections is generally extremely high. A single S. mansoni
worm pair in a mouse may be equivalent to more than 1,000 pairs in an infected
person (Cheever 1969, Gryseels & de Vlas 1996).
B. Most schistosome infections in
humans are acquired gradually over years while most experimental infections
are given as a single exposure. When a mouse is given multiple inocula, the
cumulative intensity of infection becomes progressively less realistic.
C. Most humans exposed to schistosomiasis
are born to mothers who are or have been infected. There are ample reasons to
think that in utero exposure to schistosome antigens or to idiotypic or anti-idiotypic
antibodies may modify the response to subsequent infection in humans (Eloi-Santos
et al. 1989, Novato-Silva et al. 1992) and in mice (Montesano et. al. 1997,
1999a, b).
D. The chronicity of schistosome
infections in humans is obviously not reproducible in most animal models. Although
one may predict that in some respects a week or two in the life of a mouse may
be equivalent to a year in humans, the calculation of equivalent times is uncertain.
E. While different strains (isolates)
of S. mansoni clearly behave differently in laboratory hosts it is unclear
if these patterns are relevant to human infections. Strains from patients with
acute schistosomiasis or hepatosplenic disease did not differ in the pathology
they produced in mice (Costa & Katz 1982, Costa et al. 1984) but Thiongo
et al. (1997) found differences in egg production and egg passage in the feces
of mice infected with different Kenyan strains of S. mansoni and felt
these might be related to clinical differences in infected humans.
However, a given isolate may produce
one pattern of infection (e.g. a higher proportion of eggs in the liver) in
mice (Anderson & Cheever 1972) and a different pattern in monkeys (Powers
& Cheever 1972). Rapid changes in isoenzyme patterns occur during successive
generations of newly isolated S. mansoni strains in mice so that it is
clear that genetic selection may occur rapidly in the laboratory (LoVerde et
al. 1985, Bremond et al. 1993). Pinto et al. (1997) recently documented the
greater genetic variability in worms from field isolates compared to the LE
strain long maintained in the laboratory. Passage in the molluscan host may
also result in genetic selection of the worms (Richards & Shade 1987).
ACUTE TOXEMIC SCHISTOSOMIASIS
Humans infected for the first time
with S. mansoni often develop an acute disease characterized by fever,
malaise, diarrhea, intense eosinophilia and occasionally allergic manifestations
such as asthma or angioedema (hives). Symptoms may appear before the onset of
oviposition and are accentuated after oviposition. Acute disease is virtually
unknown in residents of endemic areas but is frequent in outsiders exposed for
the first time. Acute toxemic schistosomiasis is associated with high levels
of immune complexes and with a vigorous cellular response to schistosome antigens
(Hiatt et al. 1980). Symptoms, signs and immune reactivity decrease over a period
of months while the infection continues unabated. An acute toxemic phase is
obvious in many animal models judging from the appearance of the animals and
from an initial vigorous cellular immunity and high eosinophilia. As the infection
becomes "chronic", at 10-20 wks, the appearance of the animals improves
and the cellular response to antigen is downregulated while egg laying by the
parasite continues unchanged (Tawfik et al. 1986, Damian et al.1992). Particular
efforts to study the toxemic phase have been made in baboons, which exhibit
fever as well as the other features noted in mice and other animal models (Damian
et al. 1992, 1996). It is unclear, however, how to relate the experimental acute
disease to that in humans. The cytokine patterns which may be related to the
fever, malaise and other symptoms and signs of the acute phase in baboons included
TNF-a, IL-1 and IL-6 (Damian et al. 1996), findings
remarkably similar to those later reported in humans (Jesus et al. 2002).
Decreased levels of corticotropin-releasing
hormone, adrenocorticotropic hormone and dehydroepiandroste-rone were reported
by Morales-Montor et al. (2001) in acutely infected baboons and mice but not
in rechallenged chronically infected baboons. In baboons, but not mice, the
lower hormone levels correlated with unmodulated granulomas.
THE FORMATION OF CIRCUMOVAL GRANULOMAS
AND MODULATION OF GRANULOMA SIZE
Studies in infected mice often give
different results than those obtained from intravenous injection of eggs. A
general overview will be presented here and differences between the lung model
and the use of infected mice will be detailed later.
It has long been clear that T helper
cells (CD4+ T cells) are instrumental for the formation of granulomas around
S. mansoni eggs (Mathew & Boros 1986) and that CD8+ T cells appear
to be important for downregulation of granuloma size in chronic infections (Chensue
& Boros 1979, Henderson et al. 1992), but antibody is necessary in addition
to downregulated T cells (Jankovic et al. 1998).
In acute infections (8 wk) regulation
of granuloma size seems to differ fundamentally from downregulation in chronic
infection. Thus in acute infection granuloma size seems controlled principally
by T cells and volume is decreased by IL-10 (Flores Villanueva et al. 1996),
increased in the absence of Il-10 (Wynn et al. 1998) and increased by administration
of cyclophosphamide or cymetidine (Weinstock & Boros 1981, Weinstock et
al. 1983). Cyclophosphamide also increased granuloma size in chronic infections
and one needs to postulate that the effect on T cells can partially overcome
the effects of immunoglobulins (Colley et al. 1979, Weinstock et al. 1983).
Antibody has a significant but modest effect in acute infections (Jankovic et
al. 1998).
Delayed type hypersensitivity (DTH)
is commonly taken to be indicative of a Th1-type cellular reaction, although
there are several instances in which contact hypersensitivity etc. have been
shown to be predominantly a Th2-type response (Assherson et al. 1996). The reaction
to schistosome eggs is a cell-mediated hypersensitivity reaction which has usually
been considered to be a DTH reaction. We think that the evidence indicates that
the reaction to schistosome eggs is predominantly Th2 and propose that the schistosome
granuloma be considered as a type-2 DTH, although it is clear that the granuloma
can begin as a Th1-type response and can also later be manipulated toward Th1.
Rakasz et al (1998) found that granulomas contained numerous activated Th1 and
Th0 cells but that these were under tight control.
Lenzi et al. (1998) have given a
detailed morphogenic and biomechanical description of granuloma development
and involution and have detailed the spatial deployment of collagen fibers within
the granulomas (Lenzi et al. 1999).
The Role of Cytokines in Granuloma
Formation and Downregulation (See Tables IV,
V, VI)
Several comprehensive reviews dealing
with this subject have been published recently (Lukacs & Boros 1993, Wynn
& Cheever 1995, Cheever & Yap 1997, Cheever et al. 1998, Fallon 2000)
and the discussion here will be oriented toward the models described. Adhesion
molecules such as ICAM-1 are presumably important for circulating cells to reach
the site of the granuloma (Langley & Boros 1995, Jacobs et al. 1997b) and
adhesins are upregulated in acute and chronic murine infections.
The earliest hepatic granulomas form
in a Th1 environment with downregulation of Th1 and upregulation of Th2 responses
6 wk after infection (Todt et al. 2000). This sequence is similar to that seen
after injecting eggs into the lungs (Wynn et al. 1993). The intense blood and
tissue eosinophilia and high IgE antibody levels associated with schistosome
infections suggest a Th2-type reaction. Treatment of injected or infected mice
with IFN-g results in decreased granuloma size and
hepatic fibrosis (Czaja et al. 1989a) while chronically infected (immunologically
downregulated) mice treated with IL-4 make larger granulomas than do untreated
animals (Yamashita & Boros 1992). IL-4 and IL-13 are largely compensate
for each other for formation of hepatic granulomas in infected mice. Thus only
minute granulomas are formed when both are suppressed, as in IL-4 receptor ko
mice (Jankovic et al. 1999) or IL-4 ko mice in which IL-13 action is suppressed
(Chiramonte et al. 1999b). Kaplan et al. (1998) found that Stat6 knockout mice
mounted a minimal Th2 response and formed small granulomas. Stat4 deficient
mice showed a minimal Th1 response but normal hepatic granulomas. Wynn et al.
(1995a) found that immunization with SEA and IL-12 produced immune deviation
toward Th-1 type reactions with reduction of granuloma sized. All of these effects
are consistent with the concept of the granuloma as a Th2 dominant reaction,
but under some conditions Th1 granulomatous responses may be predominant and
damaging (Stadecker & Hernandez 1998, Chen & Boros 1999, 2001, Hoffmann
et al. 2000, Rutitzkky et al. 2001). cDNA microarrays are a powerful tool for
examining arrays of activated genes in schistosome infected-mice with Th1 and
Th2 type mediated reactions and led to recognition of the importance of neutrophils
in hepatic lesions of wild type and immune deviated mice (Hoffmann et al. 2001).
In chronic infections (20 wk) Th2
responses are blunted (Grzych et al. 1991, Henderson et al. 1992, Chensue et
al. 1992) and Borojovic (1992) regards the chronic phase of murine schistosomiasis
as predominantly Th1 mediated, largely on the basis of increasing ratio of IgG2a
to IgG1 and decreasing eosinophil and IgE levels in chronically infected mice.
IFN-g, IL-4 and IL-10 exert cross-regulatory effects
on the Th1-Th2 balance as IL-4 drives the reaction toward Th2, IFN-g
toward Th1 and IL-10 may inhibit either trend depending on the circumstances
(Chensue et al. 1994a, b, Jankovic & Sher 1996, Wynn et al. 1997, 1998,
Boros & Whitfield 1998). IL-10 ko (knockout) mice also formed very large
granulomas 8 wk after infection but subsequently downregulated granuloma size
(Wynn et al. 1998).
IL-5 does not seem to be directly
involved in mediating granuloma size or fibrosis. Anti-IL-5 treated mice lacked
eosinophils but granuloma size and hepatic fibrosis were virtually unaffected
in both acute (Sher et al. 1990) and chronic (Cheever et al. 1992c) infections
and the pathology was not greatly changed in IL-5 ko mice (Rosa Brunet et al.
1999a). Rumbley et al. (1999), however, note that eosinophils form the majority
of cytokine producing cells in the granuloma and are the dominant source of
IL-4.
The use of T cell lines and clones
would seem to be a definitive way of resolving the relative importance of Th1
and Th2 cells. However granulomas are induced by transfer of Th0, Th1 and Th2
cells specific for SEA (Chickunguwo et al. 1991, Zhu et al. 1994, Jankovic et
al. unpublished).
The downregulation of granuloma size
has often been regarded as an effect of CD8+ suppressor cells (Chensue &
Boros 1979, Green & Colley 1981) which one might expect to be mediated at
least in part by IFN-g. Normal downregulation was
found in ko mice unable to make IFN-g, in CD8 ko
mice, in b-2 microglobulin ko mice (also virtually
unable to produce CD8+ cells) and in TAP1 ko mice which are unable to process
antigen in the context of class I (Hernandez et al. 1997b, Yap et al. 1997).
It is unclear how these findings can be reconciled with the studies implicating
CD8+ cells in downregulation of granuloma size. Rumbley et al. (1998, 2001)
hypothesize that selective apoptosis of sensitized lymphocytes within the granulomas
contributes to immunoregulation (Rumbley et al. 1998, 2001, Lundy et al. 2001,
Lundy & Boros 2002).
Infected mice rendered B cell deficient
by treatment with anti-µ antiserum (Cheever et al. 1985) or by genetic
manipulation (Jankovic et al. 1998, Ferru et al. 1998) (µMT ko mice) did
not modulate granuloma size or hepatic fibrosis and showed increased granuloma
size and fibrosis in acute (8 wk) and chronic infections. T cell responses to
antigen, however, were downregulated normally in chronically infected µMT
ko mice. Another type of B cell ko mouse (JHD B-less) showed no difference in
granuloma size from controls 8 wk after infection (Hernandez et al. 1997a) but
chronic infections were not examined. Mice lacking the Fc receptor also do not
modulate granuloma size normally, indicating that the effect of antibody in
modulation is directed through the Fc receptor (Jankovic et al. 1998). This
result is consistent with the reports of downregulation of in vitro granuloma
size by immune complexes (Goes et al. 1991,1994, Rezende et al. 1997b, c).
SCID mice show almost no reaction
to S. mansoni eggs but after injection with recombinant TNF-a
formed granulomas around S. mansoni eggs (Amiri et al. 1992) and infected,
immunologically intact mice treated with anti-TNF-a
formed granulomas reduced in size (Joseph & Boros 1993). Administration
of recombinant murine TNF-a to mice with chronic
S. mansoni infection restored granuloma size to that seen in acutely
infected animals (Joseph & Boros 1993). Nevertheless mice lacking both the
p55 and p 75 chains of the TNF-a receptor formed
normal granulomas in 8 wk infections (Yap et al. unpublished). Cheever et al.
(1999) were unable to affect granulomas in infected SCID mice by injection of
TNF-a.
Flores Villanueva et al. (1994) consider
the granuloma to be a Th1-type response and attribute downregulation to anergy.
Treatment of acutely infected animals with IL-10Fc fusion protein decreased
the size of granulomas in acute infection while diminishing levels of Th1 cytokines
(IFN-g and IL-2) and augmenting levels of Th2 cytokines
(Il-4 and IL-10) (Flores Villanueva et al. 1996). Chen and Boros (1998, 1999)
produced "Th1" or "Th2" granulomas to egg antigen epitopes
p38 or P4 depending on the adjuvant used in mice of the H2k haplotype.
IL-4 knockout mice exhibited increased
Th1-like responses and markedly diminished Th2 responses but granuloma size
and hepatic fibrosis during infection were equivalent to those in intact controls
(Pearce et al. 1996, Rosa Brunet et al. 1997). However, T cell responses were
more generally affected in IL-4 ko mice (Pedras-Vasconcelos et al. 2001). Metwali
et al. (1996) found granuloma size to be diminished in IL-4 ko mice but gave
a different interpretation to cytokine patterns similar to those reported by
Pearce et al. (1996) and Chiramonte et al. (1999b) found enlarged granulomas
but decreased fibrosis in IL-4 ko mice. Other experiments in various strains
of IL-4 ko mice have given discrepant results which we cannot yet interpret
(Cheever et al. 1998, Jankovic et al. unpublished).
Weinstock and his colleagues (1992,
1998) have extensively investigated the role of angiotensin converting enzyme
and neuropeptides in the formation and modulation of schistosome granulomas
(Tables IV, V,
VI). Chemokine expression has been examined
broadly by Park et al. (2001) and Qiu et al. (2001).
Role of Murine Host Genotype on
Immune Reactivity, Granuloma Size and Hepatic Fibrosis
Claas and Deelder (1979) found C3H/Sn
(H-2k) mice had lower mortality, antibody titers and DTH (ear swelling)
than did C3H.B10 (H-2b) mice but worm numbers and cell proliferation
in vitro did not differ. The differences extended to the I-region of the H-2
complex and B10.A(2R) showed lower mortality but higher antibody titers than
B10.A(4R) mice (Class & Deelder 1980). Mendlovic et al. (1989a, b) further
explored the effects of I-A and I-E on the responses of congenic mice to crude
and purified antigens.
Jones et al. (1983) used H-2b
and H-2k mice on both the C57BL/10 and BALB/c backgrounds and found
that both the strain background and the H-2 haplotype influenced antibody response
and passage of eggs in the feces. The effects of crosses between mouse strains
were complex (Jones & Kussel 1985).
Kee et al. (1986) tested fractionated
antigens in mice with a variety of haplotypes and found that a 14kDa fraction
was recognized only by H-2k mice while an 86kDa fraction was not
recognized by any of 4 H-2b strains of mice. Responses to the 86
kDa fraction were further examined by Schweitzer and Taylor (1991) and Schweitzer
(1992). Trzyna and Cordingley (1993) found H-2b mice unable to produce
IgG to a major egg antigen, p-40 and Hernandez et al. (1997c, 1998). Stadecker
and Hernandez (1998) and Hernandez and Stadecker (1999) found that H-2k
mice, including C3H and B10.Br, made a T-cell response to a recombinant fraction
of p40 while C57BL/6 mice (H-2b) did not. Granuloma size and other
pathologic changes did not, however, follow H-2 haplotypes in a large number
of mouse strains examined nor in congenic mice (Fanning et al. 1981, Colley
et al. 1983, Fanning & Kazura 1985, Cheever et al. 1987).
Comparison of Granuloma Formation
and Regulation in Infected Animals Versus the Response to Injected Eggs
The lung model allows study of granuloma
formation and cytokine production in vivo in naive animals at defined periods
after egg injection and the model allows sensitization with defined antigens
or antigenic fractions as well as the testing of defined antigens as targets,
i.e. the injection of antigens on beads which are roughly the size of schistosome
eggs. Although the lung model generally seems to reflect the sequence of events,
and regulation, present during the course of infection induced by cercariae,
important exceptions exist. Administration of IL-13R fusion protein, inhibiting
IL-13 function, dramatically reduced granuloma size around eggs injected into
the lung (Chiramonte et al. 1999a) but did not affect the size of hepatic granulomas
in infected mice (Chiramonte 1999b). Additionally, although granulomas around
injected eggs were downregulated by the injection of anti-IL-4 granulomas around
eggs laid by worms in infected animals, in the lung or in the liver, were not
(Cheever et al. 1994c). The differences were attributed to the inferior antigenic
quality of eggs extracted from the tissues as neither the site of granuloma
formation nor the state of sensitization of the host were important (Eltoum
et al. 1995). Granulomas around eggs injected into the lungs downregulate in
a fashion similar to hepatic granulomas while granulomas around pulmonary eggs
shunted from the portal system in infected mice are not downregulated (Sousa
Vidal et al. 1993). In infected IL-10 ko mice, liver granulomas were larger
than in control mice while granulomas in the lung of sensitized mice were smaller
in IL-10 ko mice than in controls (Wynn et al. 1997, 1998).
Although Eltoum et al. (1995) did
not find the site (lung vs liver) of the granulomas to be important, there are
obvious differences in the liver and lung, e.g. the hepatic and pulmonary microenvironments
are different and the miracidia in eggs injected via the tail vein are killed
much more rapidly than those in eggs injected into the portal vein (Feldman
et al. 1990) and all remnants of the eggs are removed from the lungs more quickly
(Cheever & Anderson 1971, Almeida & Andrade 1983). Kupffer cells in
the liver of infected mice contribute to the type 2 reaction (Hayashi et al.
1999).
ICAM-1 was the only adhesion molecule
identified in the liver of infected wild type (control) mice (Ritter & McKerrow
1996) while VCAM-1 was the predominant adhesion protein identified in the vicinity
of lung granulomas (Ritter & McKerrow person. commun.). VCAM-1 was seen
in the livers of infected ICAM-1 ko mice, affording a striking illustration
of the ability of ko mice to adapt to their deficiencies (Ritter & McKerrow
1996). However, VCAM-1 was found to be rapidly upregulated in the livers of
infected-wild type mice by Rathore et al. (1996), a result in apparent conflict
with the above. Leptak and McKerrow (1997) found little IL-4 in livers of mice
given injected portal eggs and postulate that TNF-a
plays a prominent role in the formation of hepatic granulomas while IL-4 is
important for initiation of pulmonary granulomas, but Eltoum et al. (1995) found
similar levels of IL-4 mRNA after injection of eggs at either site.
The regulation of granuloma size
in the mouse gut also differs from that in the liver (Weinstock & Boros
1981, Jacobs et al. 1998a).
HEPATIC FIBROSIS
General
Hepatic fibrosis is related to the
immune response to the egg and is virtually absent in infected nude and SCID
mice (Cheever et al. 1994a, Cheever et al. 1999) or in mice infected with only
male or female parasites.
Female C3H/HeNN mice infected with
25 male worms for 20 wk had only 2.21 µmoles of hydroxyproline per liver
as compared to 1.53 in uninfected mice and 30 in mice infected with 2-3 worm
pairs. However mice unisexually infected for 8 wk and then given bisexual infections
modulated fibrosis (but not granuloma size) and had 21% less fibrosis (p <
0.05 in each of two experiments) than control mice given the same infection
(Cheever et al. unpublished).
The regulation of fibrosis is often
independent of the regulation of granuloma size, i.e. larger granulomas are
not always associated with greater hepatic fibrosis (Olds et al. 1989, Cheever
et al. 1994c, Phillips et al. 1996, Cheever 1997, Chiramonte et al. 1999b, Fallon
et al. 2000b). Hepatic fibrosis in schistosome infected mice is clearly linked
to IL-13 and the Th2 response, although fibrosis also clearly sometimes occurs
around granulomas formed in the Th1 melieu (Chen & Boros 1999, Hernandez
et al. 1999, Hoffmann et al. 2000, Hesse et al. 2000).
Animals treated with anti-IL-4 showed
a decrease in Th2 response, an increased Th1 response and a reduction in hepatic
fibrosis (Cheever et al. 1994c). IL-13 is even more important for fibrosis and
treatment of infected mice with sIL-13R a2Fc fusion protein led to a marked
decrease in fibrosis with little effect on granuloma size (Chiramonte et al.
1999b). IL-4 ko mice treated with sIL-13R had tiny granulomas but fibrosis equivalent
to that in SIL-13R treated wild type (WT) mice, although the cytokine pattern
was Th2-like in WT mice and Th1-like in IL-4 ko mice (Chiramonte et al. 1999b)
and in IL-4R ko mice, in which neither IL-4 or IL-13 can signal in the absence
of the receptor (Jankovic et al. 1999). Mice treated with anti-IL-4 might be
expected to have low IL-13 levels. IL-13 ko mice had granulomas the same volume
as wild-type mice but showed minimal fibrosis (Fallon et al. 2000b). Arginase-1
activity is important in influencing both granuloma size and hepatic fibrosis,
probably through increasing hepatic proline levels (Hesse et al. 2001).
Mice vaccinated with eggs + IL-12
before infection developed an increased Th1-like cytokine response, a moderate
decrease in granuloma size and a marked decrease in hepatic fibrosis (Wynn et
al. 1995). This effect is dependent on IFN-g, IL-12
and TNF-a (Hoffmann et al. 1999) and mediated through
nitric oxide synthase-2 (Hesse et al. 2000). Treatment with IFN-g
resulted in a marked decrease in hepatic fibrosis (Czaja et al. 1989a) but anti-IFN-g
had no effect on granuloma size or hepatic fibrosis in infected mice (Sher et
al. 1990). SEA given without adjuvant intravenously or intraperitoneally before
infection also affected granuloma size (Botros et al. 1997) and fibrosis (uncorrected
for egg number, Pancré et al. 1999). Hassanein et al. (2001) noted amelioration
of pathology in mice immunized intravenously with either SEA or recombinant
S. mansoni glutathione S-transferase.
Wolowczuk et al. (1997) reported
that a single subcutaneous dose of IL-7 given at the site of exposure to S.
mansoni increased worm recovery and hepatic collagen (µg collagen/mg
protein) but their analysis of fibrosis did not allow for the increased intensity
of infection in treated mice. IL-7 ko mice had small granulomas and little fibrosis
(Woloczuk et al. 1999) and IL-7 Tg mice showed increased fibrosis (Roye et al.
2001). A somatostatin inhibitor, octreotide, diminishes hepatic fibrosis in
infected mice (Mansy et al. 1998).
Hepatic fibrosis in infected mice
is related to egg numbers, but not in a linear fashion, i.e. mice with heavier
infection have more total hepatic fibrosis but less fibrosis per egg (Cheever
1986). The rate of hepatic fibrosis, as indicated by collagen mRNA (Wynn et
al. 1995a, b) or collagen synthetic rates (Olds et al. 1989), was decreased
in chronically infected mice compared to those with acute (6-8 wk) infections.
However Takahashi and Simpser (1981) felt that decreasing levels of collagenase
in chronic infections were more important than decreased synthetic rates and
in acute infections mice receiving a single injection of concanavalin A showed
increased collagenase activity and decreased hepatic collagen (Takahashi &
Koyayashi 1982). We are unaware of attempts to adjust the rate measurements
of hepatic fibrosis for infection intensity. Collagen synthetic rates were proportional
to liver collagen per egg in ICR mice with high fibrosis compared to C57BL/6
mice with low fibrosis (Cheever et al. 1983).
Widely different rates of resorption
of hepatic collagen have been reported following chemotherapeutic cure of infection
(Morcos et al. 1983, Andrade & Grimaud 1986, Cheever et al. 1992a), but
in general collagen resorption is relatively rapid (collagen half-life of wk
or months) in mice treated during the first few weeks of egg deposition and
much more gradual when chronically infected mice were treated (Warren &
Klein 1969). The morphologic pattern of collagen resorption in mice resembles
that in humans (Andrade & Grimaud 1988, Andrade et al. 1992). Colchicine
treatment did not decrease hepatic fibrosis in S. mansoni-infected mice
(Badawy et al. 1999).
Symmers' Clay Pipestem Fibrosis
Symmers' fibrosis is present in virtually
all cases of portal hypertension attributable to schistosome infection in humans
and nearly all cases with Symmers' fibrosis at autopsy also manifest signs of
portal hypertension. Symmers' fibrosis in the chimpanzee was morphologically
indistinguishable from that in man but did not produce portal hypertension (Sadun
et al. 1970), probably because of the extensive portal-systemic collaterals
formed. Njenga et al. (1998) and Nyindo and Farah (1999) describe, but do not
convincingly illustrate, Symmers'-like periportal fibrosis in S. mansoni-infected
baboons and Farah et al. (2000) relate portal fibrosis in baboons to reinfection
and TGF-b and IL-4 production.
The massive infections present in
mice produce a portal hypertension related to granuloma number and size which
is probably not relevant to the mechanism of portal hypertension in humans (Cheever
1965a). However, mice also share the obstructive portal-venous lesions apparently
responsible for portal hypertension in humans. Portal fibrosis resembling Symmers'
fibrosis in humans was first described in mice by Warren (1966). Andrade (1987)
and Andrade et al. (1997) described similar fibrosis in infected mice in which
low intensity infections allowed gradual obstruction of peripheral portal venules
with subsequent preferential shunting of eggs into the portal spaces surrounding
larger veins. In infected persons it was also noted that eggs did not concentrate
in the large portal spaces until pipestem fibrosis had already begun (Cheever
1969). Splenectomized mice developed pipestem fibrosis but less frequently than
intact mice (Andrade et al. 1998).
Henderson et al. (1993) not only
described a more marked and uniform Symmers'-like fibrosis in infected CBA male
mice but also related this fibrosis to the absence of regulatory idiotypic antibodies
in these mice, a finding strikingly similar to their findings in humans with
Symmers' fibrosis (Montesano et al. 1990a, b). Exposure of neonatal mice to
cross-reactive idiotypes (CRI) of mice without portal fibrosis, but not to CRI
of mice with portal fibrosis, resulted in decreased granuloma size and decreased
hepatic fibrosis in mice subsequently infected (Montesano et al. 1999b). Thus
both mechanistic and immunological mechanisms in the murine model of pipestem
fibrosis resemble those in humans. These CBA mice also had elevated serum levels
of TNF-a and TNF-a mRNA
in their livers (Adewusi et al. 1996) and decreased levels of IL-10 (Bosshardt
et al. 1997). However idiotypes from mice without portal fibrosis stimulated
IFN-g formation (Montesano et al. 1997). Clinical
hepatosplenic disease in humans was associated with a Th1-type immunological
response (Mwantha et al. 1998) but Montenegro et al. (1999) saw IFN-g
production in such patients only after neutralization of IL-10. Mice with and
without pipestem hepatic fibrosis did not show differences in the production
of anti-idiotypic antibodies against S. mansoni (Andrade et al. 1998).
Gross examination of the livers of
mice (and other rodent hosts) can be misleading. Macroscopic white thickening
of the portal areas is frequent in chronic infections, but microscopically this
usually reflects marked proliferation of bile ducts rather than fibrosis. Bile
duct proliferation is often associated with eosinophilic crystals in the duct
lumen. The crystals are apparently derived from epithelial secretions and not
eosinophils. Similar changes are seen in the lungs of chronically infected mice
and are presumably IL-4 and IL-13 dependent as similar crystals are seen in
the lungs of IL-4 and IL-13 transgenic mice.
Extrahepatic Pathology
Lung- The key pulmonary pathology
in infected humans is granulomatous schistosomal pulmonary arteritis resulting
in pulmonary hypertension and cor pulmonale. Granulomatous pulmonary arteritis
is seen routinely in mice injected with schistosome eggs, but the attempts to
model human pulmonary schistosomiasis have generally involved the shunting of
eggs in infected animals to the lungs through partial ligation of the portal
vein which created a shunt from the portal system to the lungs (Warren 1964)
producing granulomatous pulmonary endarteritis similar to that in infected humans
(Andrade & Andrade 1970), lesions only partially reversed by chemotherapy
(Almeida & Andrade 1983). Granulomas in the lungs of shunted mice did not
undergo the downmodulation in size seen in the livers of the same animals after
chronic infection (Souza Vidal et al. 1993).
Pulmonary granulomas in mice with
portacaval shunts were resistant to downregulation by anti-IL-4 while granulomas
around intravenously injected eggs were downregulated, a difference attributed
to the lower antigenic potency of the eggs injected compared to those laid in
situ by the worms (Eltoum et al. 1995).
Intestines - Granulomas in
the colon and small intestine of mice are smaller than those in the liver, are
not always subject to the same downregulation (Weinstock & Boros 1981, Jacobs
et al. 1998a) and are associated with less fibrosis than are hepatic granulomas
(Dunn & Kelley 1979, Santos et al. 1992). Inflammatory colonic polyps were
seen in S. mansoni-infected chimpanzees (Sadun et al. 1970) but these
did not ulcerate and were not associated with diarrhea, a hallmark of schistosomal
colonic polyposis in man. Heavily infected animals of several species may develop
colonic ulcerations and bloody diarrhea but it is unclear whether these bear
any relation to lesions or symptoms and signs in humans. Infected woodchucks
have only slight hepatic fibrosis but marked fibrosis of the intestine (Andrade
et al. 2001).
Alterations in the vasculature and
innervation of the intestines of infected mice have been reported (Kloetzel
1971, Block 1980, Varilek et al. 1991). Death of intestinal neurons is uncommon
(Nassauw et al. 2001). In spite of the predominance of small intestinal over
colonic pathology in infected mice and the evident gross and microscopic lesions,
the functional changes seem slight. Domingo and Warren (1966) did not find functional
changes in the guts of infected mice while Vengesa and Lesse (1979) and Sadek
et al. (1986) noted changes in the absorption of glucose and fluid transport
and of disaccharidase activity. Moreels et al. (2001) found that at 12 but not
8 wk after infection that increased muscular contractility was present in the
inflammed mouse gut and that transit through the GI tract was decreased. Immunodeficient
mice may have severe intestinal disease, as noted below under "Causes of
Death".
Kidneys - The glomerulonephritis
seen in schistosome infected humans has not been described in S. mansoni
infected animals although both chimpanzees and rabbits seem susceptible to this
lesion when infected with S. japonicum. S. mansoni infected rabbits
show similar changes, albeit of a lesser degree (Andrade et al. 1988). Mild
glomerular lesions in infected capuchin monkeys were described by de Brito et
al. (1971) and in baboons by Houba et al. (1977). Houba (1979) reviewed experimental
schistosomal glomerulopathy.
Glomerular deposits of immune complexes
and ultrastructural glomerular lesions and Ig deposits have been described in
infected mice, but the kidneys were often normal by light microscopy (Andrade
& Susin 1974, Rousse & Romeiro 1974, Natali & Cioli 1976, Carneiro
& Lopes 1986, Water et al. 1988). Antigen and immune complex deposition
were augmented by portocaval shunting (van Marck et al. 1977) but were present
even in unisexual infections (Lopes et al. 1981). El-Sherif and Befus (1988)
found IgA to be the predominant Ig isotype in the glomeruli of infected mice.
Severe glomerulonephritis developed in female BXSB mice exposed to 10 S.
mansoni cercariae but most mice did not have granulomas in the liver and
unisexual infections were not ruled out (Fugiwara et al. 1988). Hematuria was
documented in a substantial proportion of infected mice by Valadares and Pereira
(1983).
S. mansoni-infected hamsters
developed severe amyloidosis of the kidneys which resulted in marked ascites
and amyloidosis of the liver and spleen, which complicates interpretation of
the lesions in these organs (Cheever 1965b).
Central Nervous System -
No proper model has been described for the focal mass lesions most frequently
reported in humans, but Aloe et al. (1996) have described granulomas in the
brain and altered nerve growth factor levels in chronically infected mice. Behavioral
changes have been noted in mice and are perhaps attributable to effects of cytokines
on the central nervous system (Fiore et al. 1996). Focal egg deposition and
granulomatous encephalitis associated with convulsions were seen in an infected
cercopithecus monkey (Cheever & Duvall 1974). Recently, Silva et al. (2002)
have called attention to the inadequacy of the murine model for studies concerning
neuroschistosomiasis.
Causes of Death
Causes of death in patients -
Humans rarely die from acute S. mansoni infections. Nearly all persons
with lethal chronic infections have Symmers' fibrosis and most of these die
from bleeding esophageal varices. Others with Symmers' fibrosis die with chronic
hepatocellular failure and a smaller number from complications of pulmonary
hypertension or glomerulonephritis (Cheever & Andrade 1967) but death from
schistosomiasis in patients without Symmers' fibrosis is very uncommon.
Variability between laboratories
- It is clear that in some laboratories lethal infections in mice are associated
with lower numbers of infecting worms than in other laboratories. It is unclear
whether this should be attributed to the parasite, the host, to concomitant
bacterial or viral infections or to other variables associated with animal husbandry.
Acute hemorrhage into the gastrointestinal
tract causes the great majority of deaths in S. mansoni infected mice
in our experience. A/J mice incur this complication early in infection and at
low infection intensities (Dean et al. 1981, Colley & Freeman 1983) while
C57BL/6 and BALB/c mice are unusually resistant to death after infection. The
mechanism of intestinal hemorrhage in mice is unrelated to gastrointestinal
hemorrhage in humans with schistosomiasis. Esophageal varices in the mouse are
on the serosal surface of the esophagus and are not the source of bleeding,
which is presumably from minute lesions in the gut. This does not seem to be
a cause of death in other models, and perhaps is lethal for the mouse because
it cannot afford to lose the 1-2ml of blood involved.
Cytokine shock in mice - Marked
malaise and early death without hemorrhage into the gut lumen were reported
in IL-4 ko mice on the C57BL/6 background, but not in IL-4 ko mice which were
F1 129JxB6. The lethal effect was related to IFN-g
and NO (La Flamme et al. 2001, Patton et al. 2002) but was independent of IL-12
(Patton et al. 2001). Survival was improved by treating the mice with anti-TNF-a
or with recombinant IL-4 (Rosa Brunet et al. 1997a, b, 1999). Mice deficient
in IL-4 and IL-13 are especially susceptible and Fallon et al. (2000b) give
an excellent detailed description of the pathogenesis of cytokine shock in these
mice, including the shift to a type 1 type cytokine response and the effects
of bacterial lipopolysaccharides leaking into the circulation from the intestine.
Fallon (2000d) provides an excellent review. Hoffmann et al. (2000) found that
either extreme Th1 or Th2 polarization resulted in the early death of S.
mansoni-infected mice but probably not from cytokine shock in Th2 polarized
mice.
Other immunocompromised mice die
in the first wk of egg laying without evident cytokine shock. Fallon et al.
(2000) found that mice transgenic for IL-9 developed a fatal enteropathy in
a Th2 melieu and MacDonald et al. (2002) noted wasting and early death of CD154
ko mice which had impaired Th2 responses.
T cell deficient mice, including
SCID and nude mice, infected with S. mansoni, but not S. japonicum
or S. haematobium, frequently die 7-9 wk after infection, apparently
from hepatotoxicity [and presumably cytokine shock] (Byram et al. 1979, Cheever
et al. 1999). Mice deficient in Class II MHC (Angyalosi et al. 1998), CD-4 depleted
mice (Fallon et al. 2000a) or made tolerant to S. mansoni egg antigens
seem to die a similar death (Fallon & Dunne 1999). Gharib et al. (1999)
have described increased eosinophil peroxidase activity and increased oxygen
radical production (Abdallahi et al. 1999) in the livers of S. mansoni-infected
mice with a concomitant decrease in hepatic antioxidant defenses, apparently
predisposing infected mice to hepatotoxicity. It is not clear that the entities
we have attributed to "cytokine shock" are all similar nor has this
toxicity in mice been shown to be relevant to death or other illness in humans
(Cheever et al. 2000). If the syndrome is related to acute schistosomiasis then
it is not clear why immunodeficient mice do not show cytokine shock when infected
with schistosome species other than S. mansoni (Cheever et al. 2000).
WORM DEVELOPMENT, FECUNDITY AND
FECAL EGG EXCRETION
Fewer worms develop in male rodent
hosts than in females and castration and exogenous hormones have the expected
effects (Berg 1957, Nakazawa et al. 1997). Fulford et al. (1998) propose that
in mice and in humans that this might be related to dehydroepiandrosterone levels.
Worm development and egg production were delayed in mice deficient in the 5th
component of complement at 7 wk of infection (Ruppel et al. 1982) but no difference
was found in another laboratory at 12 wk (Cheever et al. 1987). Biozzi high
responder mice developed higher numbers of parasites and larger worms, and significantly
more eggs/worm pair were found in the liver at 10 but not at 15 wk (Blum &
Cioli 1978). IL-7 deficient mice had fewer worms and fewer eggs/female 7-8 wk
after infection (Wolowczuk et al. 1999).
Decreased numbers of eggs/worm pair
were found in the livers of severely malnourished mice and hepatic granulomas
in these mice were smaller than in well nourished mice (Knauft & Warren
1969, Akpom & Warren 1975, Coutinho et al. 1997) and pipestem fibrosis was
absent and total hepatic fibrosis diminished in malnourished mice (Coutinho
et al. 1997).
Fecundity is defined as the number
of eggs laid per day per worm pair. Delayed inception of egg laying is more
common than decreased fecundity so that measurement of eggs/worm pair at a single
point in acute infections is of limited value. Strictly speaking one should
know the total number of eggs in the tissues and eggs passed in the feces to
calculate fecundity. Generally, only the number of eggs in the liver and intestines
is known, and this comprises almost all eggs in the tissues. Eggs destroyed
in the tissues also constitute a portion of the eggs laid and in rhesus monkeys
the rate of egg destruction was extremely rapid [half life of 8 days] (Cheever
& Powers 1971) and the same has been assumed to be true for cercopithecus
monkeys and baboons (Cheever & Duvall 1974, Damian et al. 1986). In mice
egg destruction is much slower and its effects can often be ignored (Cheever
et al. 1992a). Quantitative estimates of worm fecundity for many of these species
have been presented previously (Cheever et al. 1994b). Farah et al. (1997) described
higher fecundity in multiply infected baboons than after single infection.
In mice infected with a single pair
of worms, fecundity varied greatly between individual mice but did not vary
with duration of infection (Cheever et al. 1994a). Fecundity did decrease in
time in more heavily infected mice (Cheever et al. 1994b). Worm fecundity in
vitro also varied greatly among worm pairs and with the host used and the duration
of infection (El Ridi et al. 1997). The fecundity of worms in infected rhesus
monkeys decreased with increasing duration of infection. This decrease was much
more rapid in heavily infected animals than in those lightly infected and the
decrease in fecundity presumably had an immune basis as worms were also much
more rapidly destroyed in the heavily infected monkeys (Cheever & Powers
1969).
The fecundity of the worms was generally
reflected by the number of eggs passed in the feces at various stages of infection
in mice (Cheever et al. 1994b) and in monkeys (Cheever & Powers 1969). The
percentage of eggs laid passed in the feces can vary greatly in different species
and even in different strains of mice (Jones et al. 1989). Nude mice (Cheever
et al. 1993) and T-cell depleted mice (Dunne et al. 1983, Doenhoff et al. 1978,
1985) passed less eggs in the feces than did intact mice and SCID mice passed
almost no eggs in the feces in the first weeks of oviposition (Amiri et al.
1992, Cheever et al. 1999). These findings anticipated those in patients with
AIDS who may pass many fewer eggs in the feces than comparably infected patients
without AIDS (Karanja et al. 1997). The number of eggs per worm pair in the
tissues of nude and SCID mice was low early in infection (Amiri et al. 1992,
Cheever et al. 1993, 1999) but the rate of egg accumulation approached that
in intact mice by the 10th wk of infection (Cheever et al. 1993, 1999). Davies
et al. (2001) correlate worm development with the presence of a subset of CD4+
lymphocytes in the liver.
Lenzi et al. (1987) found that eosinophils
favored the passage of S. mansoni eggs in the feces, although mice treated
with anti-IL-5 and lacking eosinophils passed normal numbers of eggs in the
feces (Sher et al. 1990). Worms in mice deficient in IL-4 or IL-4 and IL-13
showed normal to slightly increased fecundity as judged from tissue eggs but
almost no eggs were passed in the feces (Fallon et al. 2000b).
More selective immune depletion,
i.e. treatment with anti-IgE, has been reported to inhibit the development of
S. mansoni suggesting that the worms need this immunoglobulin for normal
development (Amiri et al. 1994). This is difficult to reconcile with the increased
numbers of worms developing in IgE ko mice (King et al. 1997) and the normal
development of worms in FceRI ko mice, in B cell ko mice and in SJA/9 mice unable
to produce IgE (Jankovic et al. 1997, 1998, El Ridi et al. 1998).
INTERACTION OF SCHISTOSOMIASIS
WITH OTHER DISEASES
Murine viral hepatitis was more severe
in schistosome infected mice (Warren et al. 1969) and the clearance of vaccinia
virus was delayed for a period during the acute phase of schistosome infection
(Actor et al. 1994). Woodchuck hepatitis B infection was apparently not affected
by S. mansoni infection (Andrade et al. 2001). Surprisingly hepatitis
B replication in HBV transgenic mice was inhibited by S. mansoni infection
(McClary et al. 2000).
Schistosomiasis is intriguingly related
to several bacterial infections, most notably with prolonged typhoid fever syndromes
(Chieffi 1992) and numerous experimental studies have been published using a
number of bacterial species (Rocha et al. 1980). The association of bacteria
with the schistosome gut or tegument (Loverde et al. 1980) protects the bacteria
from chemotherapy and presumably from immune attack as well.
S. mansoni infected mice also
showed increased susceptibility to Entamoeba histolytica (Knight &
Warren 1973), Trypanosoma cruzi (Kloetzel et al. 1973) and Toxoplasma
gondii (Kloetzel et al. 1977). Marshall et al. (1999) found increased plasma
TNF-a and increased hepatotoxicity in mice with combined
S. mansoni and T. gondii infections and this effect was diminished
in IL-12 ko mice (Araujo et al. 2001). The susceptibility to Plasmodium yoelli
and P. berghei was little affected (Lwin et al. 1982).
There has long been controversy about
the association of human hepatocellular carcinomas and schistosome infections,
but S. mansoni infection is apparently not associated with hepatoma in
man (Cheever & Andrade 1967). Hepatomas are infrequently seen in infected
animals but infected mice given low doses of some carcinogens developed hepatomas
much more frequently than did mice given the carcinogen alone (Domingo et al.
1967, Kakizoe 1985).
The vigorous Th2 response to schistosome
infection may have a role in several of the above situations and in the response
to immunizing antigens such as diphtheria toxoid (Haseeb & Craig 1997).
Diabetes was ameliorated in NOD (non-obese diabetic) mice infected with S.
mansoni or injected with eggs (Cooket et al. 1999) and oral administration
of SEA plus insulin led to a Th2 response in NOD mice (Maron et al. 1998). Second
helminth infections may be influenced as well. Mice usually susceptible to Trichuris
muris resolved their infections when infected with S. mansoni (Curry
et al. 1995).
USE OF SCHISTOSOME-RELATED MODELS
FOR THE STUDY OF OTHER DISEASES
Many of these studies are implicit
in the discussion above. Schistosome infections have been widely used as an
example of hypersensitivity granulomas (Boros 1983) and as model systems to
examine the biochemistry and morphology of hepatic collagen formation (Silva
et al. 1990 Prakash et al. 1995, Frizell et al. 1995) or resorption, as noted
above. The interactions of the field of immunology and the study of the immunopathology
of schistosome infections have been mutually beneficial. The importance of anti-idiotypic
T cell responses in infectious disease was first noted in schistosome infected
individuals (Montesano et al. 1990a, b). The study of T helper subsets has been
extremely useful in analyzing infections with schistosomes and with other helminths
but investigations of these infections have also contributed substantially to
our knowledge of Th1 and Th2 lymphocyte subsets (Jankovic & Sher 1996, Jankovic
et al. 2000, MacDonald et al. 2002) and are relevant to understanding other
Th2 dominated reactions such as asthma. Sabin et al. (1996) recently used schistosome
eggs to demonstrate the importance of eosinophils as an early source of IL-4.
Coelom-associated lymphomyeloid tissue
(CALT or milky spots) is greatly expanded in schistosome infected mice and examination
of these animals has contributed to our knowledge of CALT (Lenzi et al. 1996).
Many other studies have added to our understanding of eosinophils, macrophages,
lymphocytes, non B, non T cells, endothelial cells etc.
SPECIAL CONSIDERATION OF THE RABBIT
AND WOOD-CHUCK
Rabbits are a host of choice for
the study of the pathology of schistosomiasis japonica. Large periovular granulomas
are formed in the liver during acute infection and typical areas of periportal
"pipestem" fibrosis appear. However, rabbits are not good hosts for
S. mansoni since adult worms are eliminated within a few wk after cercarial
exposure (Warren & Peters 1967, Andrade et al. 1988). Furthermore the eggs
do not mature in the tissues and do not appear in the stools. Thus the main
factor in the pathogenesis of schistosomiasis, the miracidium, is absent and
therefore there is no disseminated liver fibrosis, nor obstructive portal vascular
lesions and no portal hypertension or splenomegaly. Lesions in the liver are
produced by the worms rather than the eggs. The basic lesion is composed of
focal phlebosclerosis, vascular dilatation and polypoid endophlebitis. This
curious and characteristic lesion is formed by projections of chronically inflamed
subintimal connective tissue which are covered by hyperplastic endothelial cells.
At first they do not affect the general liver architecture but in severe chronic
infection some degree of portal and sepal fibrosis appear and may become prominent
in prolonged infections. In extremely severe cases the combination of fibrosis
and vascular proliferation may transform the liver into a huge angiomatous organ.
Thus focal portal vascular changes caused directly by living or dying adult
worms dominate the pathology of schistosomiasis mansoni in rabbits. Periovular
granulomas are not a feature and the fibrotic changes do not resemble human
"pipestem" fibrosis.
Woodchucks (Marmota monas
and M. marmota) are worth considering as an experimental model for schistosomiasis
because they are susceptible to both S. mansoni and to a hepatitis virus
very similar to human hepatitis virus B (Woodchuck hepatitis virus - WHV). They
are highly susceptible hosts with several peculiarities. Acute infection can
run a severe and fatal course if the cercarial load is heavy (1,000 cercariae
or more). Death in these cases is preceded by diarrhea due to intestinal lesions.
The most severely damaged organ is the intestines rather than the liver. Large
periovular granulomas are numerous in the submucosa and in the muscular coat,
the mucosa being least involved. Granulomas in the liver are smaller than those
in the intestines and are scattered. They cause some degree of portal fibrosis
which is never disseminated or systematized. The infection runs an auto-cure
course (Andrade et al. 2001).
SPECIAL CONSIDERATION OF CALOMYS
COLLOSUS
The first report of S. mansoni
infection in C. callosus was from Coelho et al. (1979). Borda (1972)
showed experimentally that it is permissive to S. mansoni.
Our histopathological observations
in C. callosus, infected by percutaneous exposure to 70 cercariae of
S. mansoni, and killed at 42, 55, 80, 90 and 160 days after infection
(six animals/point), have showed some peculiar aspects. In the liver, the granulomas
were of two types: larger, in portal spaces, and smaller ones, in the parenchyma,
which were often intravascular. The former were usually constituted by internal
(periovular) and peripheral zones, while the intraparenchymatous granulomas,
during all the studied period, were predominantly composed of large and mature
macrophages, often with abundant schistosomal pigment, less eosinophils and
neutrophils, and scarce lymphocytes, as described (Lenzi JA et al. 1995, 1998).
There was no significant change in the granuloma morphology, size and extracellular
matrix composition during the studied periods, suggesting that they are not
modulated.
The granulomas exhibited fine texture
of reticular fibers, mainly in the periphery and few or absent collagen fibers
were visible by picrosirius (Junqueira et al. 1979) or phosphomolybdic acidpicrosirius
stains for confocal laser scanning microscopy (Dolber & Sprach 1993) (Fig.
1). However, they were rich in carboxylated proteoglycans. By immunohistology,
they showed deposits of fibronectin, laminin, collagen types I, ProIII, IV,
V, which remained more or less constant during all the time of infection, always
being less than seen in mice. Gelatinase was detected in large macrophages surrounding
the eggs.
Focal parenchymal necrosis was found
and portal inflammation and fibrosis were varied, depending on the time of infection.
They were absent or minimal in more recent infection (42 days). At 90 days in
some animals, there was an exacerbation of portal inflammation, and there appeared
also portal perivenular and periductal fibrosis, causing sac-like dilatations
in portal lymphatics, which filled with large macrophages.
Although the granulomas never exhibited
peripheral metaplasia of hematopoietic cells, as observed in mice (Lenzi HL
et al. 1995), from 55 days on, intrasinusoidal megakaryocyte, eosinophil and
erythroid metaplasia was observed.
In C. callosus intestines,
there were numerous eggs in all layers forming granulomas with collagen and
reticular fibers detected only in the peripheral zone (Fig.
2). Numerous intestinal nodules, appearing from 55 ups to 160 days after
infection, were localized at the interface between external muscular layer and
intestinal serosa (Fig. 3) (Lenzi
JA et al. 2002). From 55 days of infection on, an intense mast cell infiltration
was detected, occupying all the intestinal layers. In the mucosa, the mast cells
were mainly of the mucosal mast celltype (MMC), while, in the muscular layer
and in the subserosal nodules they were almost exclusively of the connective
tissue type (CTMC) or transitional from MMC to CTMC.
Pancreatic involvement was frequent,
and occasionally severe (Figs 4,
5). The omentum and mesentery
were strongly activated during the infection, showing many milky spots and a
diffuse infiltration of mast cells (Fig.
6). Milky spots were mainly of the lymphoplasmacytic type (Lenzi et al.
1996).
ACKNOWLEDGEMENTS
To Michelle Frazier-Jessen, Dragana
Jankovic, Fred Lewis, Edward Pearce, Laura Rosa-Brunet, W Evan Secor, Sharon
Wahl, Thomas Wynn and George Yap for unpublished data and/or for their comments
on the manuscript.
REFERENCES
- Abdallahi CMS, Hanna S, DeReggi
M, Gharib B 1999. Visualization of oxygen radical production in mouse liver
in response to infection with Schistosoma mansoni. Liver 19:
495-500. [ Medline
]
- Actor JK, Marshall MA, Eltoum
IA, Buller RML, Berzofsky JA, Sher A 1994. Increased susceptibility of mice
infected with Schistosoma mansoni to recombinant vaccinia virus: association
of viral persistence with egg granuloma formation. Eur J Immunol 24:
3050-3056. [ Medline
]
- Adewusi OI, Nis NA, Lu X, Colley
DG, Secor E 1996. Schistosoma mansoni: relationship of tumor necrosis
factor-a to morbidity and collagen deposition in chronic experimental infection.
Exp Parasitol 84: 115-123. [
Medline
]
- Akhiani AA, Lycke N, Nilsson LA,
Olling S, Ouchterlony O 1996. Lack of interferon-g receptor does not influence
the outcome of infection in murine schistosomiasis mansoni. Scand J Immunol
43: 257-262. [
Medline
]
- Akpom CA, Warren KS 1975. Calorie
and protein malnutrition in chronic murine schistosomiasis mansoni: effect
on the parasite and the host. J Inf Dis 132: 6-14.
- Almeida MAC, Andrade ZA 1983.
Effect of chemotherapy on experimental pulmonary schistosomiasis. Am J
Trop Med Hyg 32: 1049-1054. [
Medline
]
- Aloe L, Moroni R, Fiore M, Angelucci
F 1996. Chronic parasite infection in mice induces brain granulomas and differentially
alters brain nerve growth-factor levels and thermal responses in paws. Acta
Neuropathol 92: 300-305.
- Amiri P, Haak-Frendscho M, Robbins
K, McKerrow JH, Stewart T, Jardieu P 1994. Anti-immunoglobulin E treatment
decreases worm burden and egg production in Schistosoma mansoni-infected
normal and interferon g knockout mice. J Exp Med 180: 43-51.
[ Medline
]
- Amiri P, Locksley RM, Parslow
TG, Sadick M, Rector E, Ritter D, McKerrow JH 1992. Tumor necrosis factor
a restores granulomas and induces parasite egg-laying in schistosome-infected
SCID mice. Nature 356: 604-607.
- Anderson LA, Cheever AW 1972.
Comparison of geographical strains of Schistosoma mansoni in the mouse.
Bull WHO 46: 233242.
- Anderson WI, King JM, Uhl EM,
Hornbuckle BC, Tenant BC 1991. Pathology of experimental Schistosoma mansoni
infection in the eastern woodchuck. Vet Pathol 28: 245-247.
[ Medline
]
- Andrade ZA 1987. Pathogenesis
of pipe-stem fibrosis of the liver. (Experimental observation on murine schistosomiasis).
Mem Inst Oswaldo Cruz 82: 325-334. [
Medline
]
- Andrade ZA, Andrade SG 1970. Pathogenesis
of schistosomal pulmonary arteritis. Am J Trop Med Hyg 19: 305-310.
[ Medline
]
- Andrade ZA, Grimaud J-A 1986.
Evolution of the schistosomal hepatic lesions in mice after curative chemotherapy.
Am J Pathol 124: 59-65. [
Medline
]
- Andrade ZA, Grimaud JA 1988. Morphology
of chronic collagen resorption. A study of the late stages of schistosomal
granuloma involution. Am J Pathol 132: 389-399. [
Medline
]
- Andrade ZA, Susin M 1974. Renal
changes in mice infected with Schistosoma mansoni. Am J Trop Med
Hyg 23: 400-403. [
Medline
]
- Andrade ZA, Warren KS 1964. Mild
prolonged schistosomiasis in mice: alterations in host response with time
and the development of portal fibrosis. Trans R Soc Trop Med Hyg 58:
53-57.
- Andrade ZA, Berthillon P, Paraná
R, Grimaud JÁ, Trepo C 2001. Schistosomiasis mansoni and viral B hepatitis
in woodchucks. J Hepatology 34: 134-139.
- Andrade ZA, Peixoto E, Gukerret
S, Grimaud JA 1992. Hepatic connective tissue changes in hepatosplenic schistosomiasis.
Hum Pathol 23: 566-573. [
Medline
]
- Andrade ZA, Ramos E, Reis MG 1988.
A patologia da esquis-tossomose mansoni no coelho. Mem Inst Oswaldo Cruz
83: 323-332.
- Andrade ZA, Silva LM, Souza MM
1997. An experimental approach to the pathogenesis of "pipestem"
fibrosis (Symmers' fibrosis of the liver). Mem Inst Oswaldo Cruz 92:
699-706. [ Medline
] [ Lilacs
] [ SciELO
]
- Andrade ZA, Silva LM, Souza MM,
Sadigursky M, Barbosa A, Oliveira IR de 1998. Role of the spleen on the pathogenesis
of schistosomal periportal (pipestem) fibrosis of the liver: an experimental
approach. Am J Trop Med Hyg 59: 557-562. [
Medline
]
- Angyalosi G, Pancré V,
Herno J, Auriault C 1998. Immunological response of major histocompatability
complex class II-deficient (Abo) mice infected by the parasite
Schistosoma mansoni. Scand J Immunol 48: 159-169.
- Araujo MI, Bliss SK, Suzuki Y,
Alcaraz A, Denkers EY, Pearce EJ 2001. Interleukin-12 promotes pathologic
liver changes and death in mice coinfected with Schistosoma mansoni
and Toxoplasma gondii. Infect Immun 69: 1454-1462.
[ Medline
]
- Assherson GL, Dieli F, Sireci
G, Salerno A 1996. Role of IL-4 in delayed type hypersensitivity. Clin
Exp Immunol 103: 1-4.
- Badawy AA, El-Badrawy NM, Hassan
MM, Ebeid FA 1999. Colchicine therapy for hepatic murine schistosomal fibrosis:
image analysis and serological study. Int J Exp Path 80: 25-34.
- Barral-Netto M, Cheever AW, Lawley
TJ, Ottesen EA 1983. Cell-mediated and humoral immune responses in capuchin
monkeys infected with Schistosoma japonicum or S. mansoni. Am
J Trop Med Hyg 32: 1335-1343. [
Medline
]
- Berg E 1957. Effects of castration
and testosterone in male mice on Schistosoma mansoni. Trans R Soc Trop
Med Hyg 51: 353-358.
- Bergquist NR, Colley DG 1998.
Schistosomiasis vaccines: research to development. Parasitol Today
14: 99-104.
- Block EH 1980. In vivo microscopy
of schistosomiasis. II. The migration of Schistosoma mansoni in the
lung, liver and intestine. Am J Trop Med Hyg 29: 62-70.
- Blum K, Cioli D 1978. Behavior
of Biozzi high and low responder mice upon infection with Schistosoma mansoni.
Eur J Immunol 8: 52-56. [
Medline
]
- Blum AM, Metwali A, Cook G, Mathew
RC, Elliott D, Weinstock JV 1993. Substance P modulates antigen-induced, IFN-g
production in murine schistosomiasis mansoni. J Immunol 151:
225-233. [ Medline
]
- Blum AM, Metwali A, Elliott D,
Li J, Sandor M, Lynch R, Weinstock JV 1997. IL-6 deficient C57BL/6 mice form
granulomas in murine schistosomiasis that exhibit an abnormal B-cell response.
J Allergy Clin Immunol 99: 74-74.
- Blum AM, Metwali A, Elliott D,
Li J, Sandor M, Weinstock JV 1998. IL-6-deficient mice form granulomas in
murine schistosomiasis that exhibit an altered B cell response. Cellular
Immunol 188: 64-72.
- Blum AM, Metwali A, Kim-Miller
M, Li J, Qadir K, Elliott DE, Lu B, Fabry Z, Gerard N, Weinstock JV 1999.
Substance P receptor is necessary for a normal granulomatous response in murine
schistosomiasis mansoni. J Immunol 162: 6080-6085. [
Medline
]
- Blum AM, Metwali A, Mathew RC,
Cook G, Elliot D, Weinstock JV 1992. Granuloma T lymphocytes in murine schistosomiasis
mansoni have somatostatin receptors and respond to somatostatin with decreased
IFN-g secretion. J Immunol 149: 3621-3626.
[ Medline
]
- Borda CE 1972. Infecção
Natural e Experimental de Alguns Roedores pelo Schistosoma mansoni Sambon,
1907, MSc Thesis, ICBUFMG, Belo Horizonte, 43 pp.
- Borojevic R 1992. Experimental
murine schistosomiasis man-soni: establishment of the chronic phase of the
disease. Mem Inst Oswaldo Cruz 87 (Suppl. V): 171-174.
- Boros DL 1983. Granulomatous inflammations.
Prog Allergy 24: 183-267.
- Boros DL, Whitfield JR 1998. Endogenous
IL-10 regulates IFN-g and IL-5 cytokine production
and the granulomatous response in schistosomiasis manoni-infected mice. Immunology
94: 481-487. [
Medline
]
- Boros DL, Whitfield JR 1999. Enhanced
Th1 and dampened Th2 responses synergize to inhibit acute granulomatous and
fibrotic responses in murine schistosomiasis mansoni. Infect Immun
67: 1187-1193. [
Medline
]
- Boros DL, Lande MA, Carrick Jr
L 1983. The Influence of host and parasite factors on Schistosoma mansoni
egg-induced tissue fibrosis. Contr Microbiol Immunol 7: 230-236.
- Botros SS, Doughty BL, Shaker
ZA, Akl MM, Sharmy R, Diab TM, Hassanein HI 1997. Efficacy of an antipathology
vaccine in murine schistosomiasis adminstered with and without chemotherapy.
Int J Immunopharmac 18: 707-718.
- Bremond P, Pasteur N, Combes C,
Renaud F, Theron A 1993. Experimental host-induced selection in Schistosoma
mansoni strains from Guadeloupe and comparison with natural observations.
Heredity 70: 33-37.
- Brito JM, Borojevic R 1997. Liver
granulomas in schistosomiasis - mast cell-dependent induction of SCF expression
in hepatic stellate cells is mediated by TNF-alpha.
J Leukocyte Biol 62: 389-396.
- Brown DR, Fowell DJ, Corry DB,
Wynn TA, Moskowitz NH, Cheever AW, Locksley RM, Reiner SL 1996. b-2-microglobulin-dependent
NK 1.1+ T cells are not essential for T helper cell 2 immune responses.
J Exp Med 184: 1295-1304. [
Medline
]
- Byram JE, Doenhoff MJ, Musallam
R, Brink LH, von Lichtenberg F 1979. Schistosoma mansoni infections
in T-cell deprived mice, and the ameliorating effect of administering homologous
chronic infection serum. II. Pathology. Am J Trop Med Hyg 28:
274-285. [ Medline
]
- Carneiro CRW, Lopes JD 1986. Surface
antigen detected by a Schistosoma mansoni monoclonal antibody in worm
extracts and kidney deposits of infected mice and hamsters. Infect Immun
52: 230-235. [
Medline
]
- Catapani WR, Parise ER, Mor MMBL,
Sadigursky M, Andrade ZA, Sant'anna OA, Misputen SJ, Siqueira M 1994. Parasite
and egg burden, hepatic collagen and histologic pattern of liver granulomas
in selection III high and low antibody responder mice infected with Schistosoma
mansoni. Mem Inst Oswaldo Cruz 89: 63-67. [
Medline
] [ Lilacs
]
- Cêtre C, Pierrot C, Cocude
C, Lafitte S, Capron A, Capron M, Khalife J 1999. Profiles of Th1 and Th2
cytokines after primary and secondary infection by Schistosoma mansoni
in the semipermissive rat host. Infect Immun 67: 2713-2719.
[ Medline
]
- Cheever AW 1965a. A comparative
study of Schistosoma mansoni infections in mice, gerbils, multimammate
rats and hamsters. I. The relation of portal hypertension to size of hepatic
granulomas. Am J Trop Med Hyg 14: 211226.
- Cheever AW 1965b. A comparative
study of Schistosoma mansoni infections in mice, gerbils, multimammate
rats and hamsters. II. Qualitative pathological differences. Am J
Trop Med Hyg 14: 227238.
- Cheever AW 1986. The intensity
of experimental schistosome infections modulates hepatic pathology. Am
J Trop Med Hyg 35: 124-133. [
Medline
]
- Cheever AW 1969. Quantitative
comparison of the intensity of Schistosoma mansoni infection in man
and experimental animals. Trans R Soc Trop Med Hyg 63: 781-795.
[ Medline
]
- Cheever AW 1997. Differential
regulation of granuloma size and hepatic fibrosis in schistosome infections.
Mem Inst Oswaldo Cruz 92: 689-692. [
Medline
] [ Lilacs
] [ SciELO
]
- Cheever AW, Anderson LA 1971.
Rate of destruction of Schistosoma mansoni eggs in the tissues of mice.
Am J Trop Med Hyg 20: 62-68. [
Medline
]
- Cheever AW, Andrade ZA 1967. Pathological
lesions associated with Schistosoma mansoni infection in man. Tran
R Soc Trop Med Hyg 61: 626-639.
- Cheever AW, Duvall RH 1974. Single
and repeated infections of grivet monkeys with Schistosoma mansoni:
parasitological and pathological observations over a 31-month period. Am
J Trop Med Hyg 23: 884-894. [
Medline
]
- Cheever AW, Powers KG 1969. Schistosoma
mansoni infection in rhesus monkeys: changes in egg production and egg
distribution in prolonged infections in intact and splenectomized monkeys.
Ann Trop Med Parasitol 63: 8393.
- Cheever AW, Powers KG 1971. Rate
of destruction of Schistosoma mansoni eggs and adult worms in the tissues
of rhesus monkeys. Am J Trop Med Hyg 20: 69-76. [
Medline
]
- Cheever AW, Yap GS 1997. Immunologic
basis of disease and disease regulation in schistosomiasis. Chem Immunol
66: 159-176. [
Medline
]
- Cheever AW, Byram JE, Hieny S,
von Lichtenberg F, Lunde ML, Sher A 1985. Immunopathology of Schistosoma
japonicum and S. mansoni infections in B cell depleted mice. Parasite
Immunol 7: 399-413.
- Cheever AW, Dunn MA, Dean DA,
Duvall RH 1983. Differences in hepatic fibrosis in ICR, C3H and C57BL/6 mice
infected with Schistosoma mansoni. Am J Trop Med Hyg 32: 1364-1369.
[ Medline
]
- Cheever AW, Duvall RH, Hallack
TA Jr, Minker RG, Malley JD, Malley KG 1987. Variation of hepatic fibrosis
and granuloma size among mouse strains infected with Schistosoma mansoni.
Am J Trop Med Hyg 37: 85-97. [
Medline
]
- Cheever AW, Eltoum IA, Andrade
ZA, Cox TM 1993. Biology and pathology of Schistosoma mansoni and S.
japonicum infection in several strains of nude mice. Am J Trop Med
Hyg 48: 496-503. [
Medline
]
- Cheever AW, Hoffmann KF, Wynn
TA 2000. Immunopathology of schistosomiasis mansoni in mice and men. Immunol
Today [letter] 21: 465-466.
- Cheever AW, Jankovic D, Yap GS,
Kullberg MC, Sher A, Wynn TA 1998. Role of cytokines in the formation and
down-regulation of hepatic circumoval granulomas and hepatic fibrosis in Schistosoma
mansoni-infected mice. Mem Inst Oswaldo Cruz 93 (Suppl.
I): 25-32.
- Cheever AW, Lewis FA, Wynn TA
1997. Schistosoma mansoni: unisexual infections sensitize mice for
granuloma formation around eggs. Parasitol Res 83: 57-59.
- Cheever AW, Macedonia JG, Deb
S, Cheever EA, Mosimann JR 1992a. Persistence of eggs and hepatic fibrosis
after treatment of Schistosoma mansoni-infected mice. Am J Trop
Med Hyg 46: 752-758. [
Medline
]
- Cheever AW, Macedonia JG, Mosimann
JE, Cheever EA 1994a. Kinetics of egg production and egg excretion by Schistosoma
mansoni and Schistosoma japonicum in mice infected with a single
pair of worms. Am J Trop Med Hyg 50: 269-280.
- Cheever AW, Mosimann JE, Deb S,
Cheever EA, Duvall RH 1994b. Natural history of Schistosoma mansoni
infection in mice: egg production, egg passage in the feces and contribution
of host and parasite death to changes in worm numbers. Am J Trop Med Hyg
50: 281-295.
- Cheever AW, Poindexter RW, Wynn
TA 1999. Egg laying is delayed but worm fecundity is normal in SCID mice infected
with Schistosoma japonicum and S. mansoni with or without recombinant
tumor necrosis alpha treatment. Infect Immun 67: 2201-2208.
[ Medline
]
- Cheever AW, Xu Y, Macedonia JG,
Cox T, Caspar P, Hieny S, Sher A 1992c. The role of cytokines in the pathogenesis
of hepatic granulomatous disease in Schistosoma mansoni infected mice.
Mem Inst Oswaldo Cruz 87 (Suppl. IV): 81-85.
- Cheever AW, Williams ME, Wynn
TA, Finkelman FD, Seder RA, Cox TM, Hieny S, Caspar P, Sher A 1994c. Anti-interleukin-4
treatment of Schistosoma mansoni-infected mice inhibits development
of T cells and non-B, non-T cells expressing Th2 cytokines while decreasing
egg-induced hepatic fibrosis. J Immunol 153: 753-759.
- Chen Y, Boros DL 1998. Identification
of the immunodominant T cell epitope of p38, a major egg antigen, and characteriztion
of the epitope-specific Th responsiveness during murine schistosomiasis mansoni.
J Immunol 160: 5420-5427. [
Medline
]
- Chen Y, Boros DL 1999. Polarization
of the immune response to single immunodominant epitope of p38, a major Schistosoma
mansoni egg antigen, generates Th1- or Th2-type cytokines and granulomas.
Infect Immun 67: 4570-4577. [
Medline
]
- Chen YG, Boros DL 2001. The Schistosoma
mansoni egg-derived r38 peptide-induced Th1 response affects the synchronous
pulmonary but not ehe asynchronous hepatic granuloma growth. Parasite Immunol
23: 43-50. [
Medline
]
- Chensue SW, Boros DL 1979. Modulation
of granulomatous hypersensitivity. I. Characterization of T lymphocytes involved
in the adoptive suppression of granuloma formation in Schistosoma mansoni-infected
mice. J Immunol 123: 1409-1414. [
Medline
]
- Chensue SW, Remick DG, Higashi
GI, Boros DI, Kunkel SL 1986. Modulation of murine schistosomiasis by exogenously
administered prostaglandins. Am J Pathol 125: 28-34.
[ Medline
]
- Chensue SW, Terebuh PD, Warmington
KS, Hershey SD, Evanoff HL, Kunkel SL, Higashi GI 1992. Role of Il-4 and IFN-g
in Schistosoma mansoni egg-induced granuloma formation. Orchestration,
relative contribution, and relationship to macrophage function. J Immunol
148: 900-906 [
Medline
]
- Chensue SW, Warmington KS, Alllenspach
EJ, Lu B, Gerard C, Kunkel SL, Lukacs NW 1999. differential expression and
cross-regulatory function of RANTES during mycobacterial (Type 1) and schistosomal
(Type 2) antigen-elicited granulomatous inflammation. J Immunol 163:
165-173. [ Medline
]
- Chensue SW, Warmington KS, Lukacs
NW, Lincoln PM, Burdick MD, Strieter RM, Kunkel S 1995a. Monocyte chemotactic
protein expression during schistosome egg granuloma formation. Am J Pathol
146: 130-138. [
Medline
]
- Chensue SW, Warmington KS, Ruth
J, Lincoln PM, Kunkel SL 1994a. Cross-regulatory role of interferon-gamma
(IFN-g), IL-4 and IL-10 in schistosome egg granuloma
formation: in vivo regulation of Th activity and inflammation. Clin
Exp immunol 98: 395-400. [
Medline
]
- Chensue SW, Warmington KS, Ruth
J, Lincoln P, Kuo M-C, Kunkel SL 1994b. Cytokine responses during mycobacterial
and schistosomal antigen-induced pulmonary granuloma formation. Am J Pathol
145: 1105-1113. [
Medline
]
- Chensue SW, Warmington KS, Ruth
JR, Lincoln P, Kunkel SL 1995b. Cytokine function during mycobacterial and
schistosomal antigen-induced pulmonary granuloma formation. J Immunol
154: 5969-5976. [
Medline
]
- Chensue SW, Warmington KS, Ruth
JH, Sanghi P, Lincoln P, Kunkel SL 1996. Role of monocyte chemoattractant
protein-1 (MCP-1) in Th1 (mycobacterial) and Th2 (schistosomal) antigen-induced
granuloma formation. J Immunol 157: 4602-4608. [
Medline
]
- Chesney J, Metz C, Stavitsky AB,
Bacher M, Bucala R 1998. Regulated production of type I collagen and inflammatory
cytokines by peripheral blood fibrocytes. J Immunol 160: 419-425.
[ Medline
]
- Chieffi PP 1992. Interrelationship
between schistosomiasis and concomitant diseases. Mem Inst Oswaldo Cruz
87 (Suppl. IV): 291-296.
- Chikunguwo SM, Kanazawa T, Dayal
Y, Stadecker MJ 1991. The cell-mediated response to schistosomal antigens
at the clonal level: in vivo function of cloned murine egg antigen-specific
CD4+ T helper type 1 lymphocytes. J Immunol 147:
3921-3925. [ Medline
]
- Chiramonte MG, Donaldson DD, Cheever
AW, Wynn TA 1999b. An IL-13 inhibitor blocks the development of hepatic fibrosis
during a t-helper type-2 dominated inflammatory response. J Clin Invest
104: 777-785.
- Chiramonte MG, Schopf LR, Neben
TY, Cheever AW, Donaldson DD, Wynn TA 1999a. IL-13 is a key regulatory cytokine
for Th2 cell-mediated pulmonary granuloma formation and IgE responses induced
by Schistosoma mansoni eggs. J Immunol 162: 920-930.
- Claas FHJ, Deelder AM 1979. H-2
linked immune response to murine experimental Schistosoma mansoni infection.
J Immunogenetics 6: 167-175.
- Class FHJ, Deelder AM 1980. Influence
of the I-region of the H-2 complex on the immune response to murine Schistosoma
mansoni infections. Acta Leidensia 48: 23-27.
- Coelho PMZ, Dias M, Mayrink W,
Magalhães P, Mello MN, Costa CA 1979. Wild reservoirs of Schistosoma
mansoni, from Caratinga, an endemic schistosomiasis area of Minas Gerais
State, Brazil. Am J Trop Med Hyg 28: 163164.
- Colley DG, Freeman Jr GI 1983.
Difference in adult Schistosoma mansoni worm burden requirements for
the establishment of resistance to reinfection in inbred mice. II. C57BL/KsJ,
SWR/J, SJL/J, BALB/cAnN, DBA/2N, AJ, B10.A(eR) and B10.A(5R) mice. Am J
Trop Med Hyg 32: 543-549.
- Colley DG, Lewis FA, Todd CW 1979.
Adoptive suppression of granuloma formation by T lymphocytes and by lymphoid
cells sensitive to cyclophosphamide. Cell Immunol 46: 192-200.
[ Medline
]
- Cooke A, Tonks P, Jones FM, O'shea
H, Hutchings P, Fulford AJC, Dunne DW 1999. Infection with Schistosoma
mansoni prevents insulin dependent diabetes mellitus in non-obese diabetic
mice. Parasite Immunol 21: 169-176. [
Medline
]
- Costa MFFL, Katz N 1982. Comparative
study of Schistosoma mansoni strains isolated from patients with toxemic
or intestinal forms of schistosomiasis. Am J Trop Med Hyg 31:
499-504. [ Medline
]
- Costa MFFL, Roche RS, Katz N 1984.
Comparative study of Schistosoma mansoni isolated from patients with
he-patosplenic and intestinal clinical forms of schistosomiasis. Am J Trop
Med Hyg 33: 918-923. [
Medline
]
- Coulson PS 1997. The radiation-attenuated
vaccine against schistosomes in animal models: paradigm for a human vaccine?
Adv Parasitol 39: 272-336.
- Coutinho EM, de Souza MM, Silva
LM, Cavalcanti CL, de Araujo RE, Barbosa Jr. AA, Cheever AW, Andrade ZA 1997.
Pathogenesis of schistosmal `pipestem' fibrosis: a low-protein diet inhibits
the development of `pipestem' fibrosis in mice. Int J Exp Pathol 78:
337-342.
- Cuison A, Tasaka K, Chuang C-K,
Minai M, Yoshikawa H, Nakajima Y 1995. Schistosome eggs in the portal vein
can induce tolerance. Int J Parasitol 25: 993-998. [
Medline
]
- Curry AJ, Else KJ, Jones F, Bancroft
A, Grencis RK, Dunne DW 1995. Evidence that cytokine-mediated immune interactions
inducted by Schistosoma mansoni alter disease outcome in mice concurrently
infected with Trichuris muris. J Exp Med 181: 769-774.
[ Medline
]
- Czaja MJ, Weiner FR, Flanders
KC, Giambrone M-A, Wind R, Biempica L, Zern MA 1989b. In vitro and in vivo
association of transforming growth factor-b1 with hepatic fibrosis. J Cell
Biol 108: 2477-2482. [
Medline
]
- Czaja MJ, Weiner FR, Takahashi
S, Giambrone M-A, van der Meide PH, Schellekens H, Biempka L, Zern MA 1989a.
g-interferon treatment inhibits collagen deposition in murine schistosomiasis.
Hepatology 10: 795-800.
- Damian RT, Chapman RW 1983. The
fecundity of Schistosoma mansoni in baboons, with evidence for a sex
ratio effect. J Parasitol 69: 987-989. [
Medline
]
- Damian RT, Rawlings CA, Bosshardt
SC 1986. The fecundity of Schistosoma mansoni in chronic nonhuman primate
infections and after transplantation into naive hosts. J Parasitol
72: 741-747. [
Medline
]
- Damian RT, Rosa MA de la, Murfin
DJ, Rawlings CA, Weina PJ, Xue YP 1992. Further development of the baboon
as a model for acute schistosomiasis. Mem Inst Oswaldo Cruz 87
(Suppl. IV): 261-269.
- Damian RT, Rosa M de la, Bucci
JG, Mohamed FM, Kanamura HY, Chou T-M, Stich RW, Rawlings CA, Weina PJ 1996.
Hypothesis derived from the baboon model for the etiology of the symptoms
of acute schistosomiasis. Am Soc Parasitologists (Abst 106).
- Davies SJ, Grogan JL, Blank RB,
Lim KC, Locksley RM, McKerrow JH 2001. Modulation of blood fluke development
in the liver by hepatic CD4+ lymphocytes. Science 294:
1358-1361. [ Medline
]
- Dean DA, Bukowski MA, Cheever
AW 1981. Relationshiop between acquired resistance, portal hypertension, and
lung granulomas in ten strains of mice infected with Schistosoma mansoni.
Am J Trop Med Hyg 30: 806-814.
- Debrito T, Gunji J, Camargo ME,
Ceravolo A, DA Silva LC 1971. Glomerular lesions in experimental infections
of Schistosoma mansoni in Cebus apella monkeys. Bull WHO
45: 419-422.
- Denoya VA, Pointer JP, Colmenares
C, Theron A, Balzan C, Cesari IM, Gonzalez S, Noya O 1997. Natural Schistosoma
mansoni infection in wild rats from Guadeloupe - parasitological and immunological
aspects. Acta Trop 68: 11-21.
- Doenhoff M, Musallam R, Bain J,
McGregor A 1978. Studies on the host-parasite relationship in Schistosoma
mansoni infected mice: the immunological dependence of parasite egg excretion.
Immunology 35: 771-778. [
Medline
]
- Doenhoff MJ, Hassounah OA, Lucas
SB 1985. Does the immunopathology induced by schistosome eggs potentiate parasite
survival? Immunol Today 6: 203-206.
- Dolber PC, Spach MS 1993. Conventional
and confocal fluorescence microscopy of collagen fibers in the heart. J
Histochem Cytochem 41: 465469.
- Domingo EO, Warren KS, Stenger
RJ 1967. Increased incidence of hepatoma in mice with chronic schistosomiasis
mansoni treated with a carcinogen. Am J Pathol 51: 307-321.
[ Medline
]
- Dunn MA, Kelley EP 1979. Organ
specificity of fibrosis in murine schistosomiasis. Clin Res 27:
265A.
- Dunne DW, Hassounah O, Musallam
R, Lucas S, Pepys MB, Baktz M, Doenhoff M 1983. Mechanisms of Schistosoma
mansoni egg excretion: parasitological observations in immunosuppressed
mice reconstituted with immune serum. Parasite Immunol 5: 1-12.
- Edungbola LD, Schiller EL 1979.
Histopathology of hepatic and pulmonary granulomata experimentally induced
with eggs of Schistosoma mansoni. J Parasitol 65: 253-261.
[ Medline
]
- Eloi-Santos SM, Novatoi-Silva
E, Maselli VM, Gazzinelli G, Colley DG, Correa-Oliveira R 1989. Idiotypic
sensitization in utero of children born to mothers with schistosomiasis or
Chagas' disease. J Clin Invest 84: 1028-1031. [
Medline
]
- El Ridi R, Ozaki T, Kamiya H 1998.
Schistosoma mansoni infection in IgE-producing and IgE-deficient mice.
J Parasitol 84: 171-174. [
Medline
]
- El Ridi R, Ozaki T, Inaba T, Ito
M, Kamiya H 1997. Schistosoma mansoni oviposition in vitro reflects
worm fecundity in vivo: individual-, parasite age- and host-dependent
variations. Int J Parasitol 27: 381-387. [
Medline
]
- El-Sherif AK, Befus D 1988. Predominance
of IgA deposits in glomeruli of Schistosoma mansoni infected mice.
Clin Exp Immunol 71: 39-44. [
Medline
]
- Eltoum IA, Wynn TA, Poindexter
RW, Finkelman FD, Lewis FA, Sher A, Cheever AW 1995. Suppressive effect of
IL-4 neutralization differs for granulomas around Schistosoma mansoni
eggs injected into mice compared to eggs laid in infected mice. Infect
Immunity 69: 2532-2536.
- Epstein MM, DiRosa F, Jankovic
D, Sher A, Matzinger P 1995. Successful T cell priming in B cell-deficient
mice. J Exp Med 182: 915-922. [
Medline
]
- Erickson DG, vonLichtenberg F,
Sadun EH, Lucia HL, Hickman RL 1971. Comparison of Schistosoma haematobium,
S. mansoni and S. japonicum in the owl monkey, Aotus trivirgatus.
J Parasitol 57: 543-558. [
Medline
]
- Fallon PF 2000. Immunopathology
of schistosomiasis: a cautionary tale of mice and men. Immunol Today
21: 29-35. [
Medline
]
- Fallon PG, Dunne DW 1999. Tolerization
of mice to Schistosoma mansoni egg antigens causes elevated Type 1
and diminished type 2 cytokine responses and increased mortality in acute
infection. J Immunol 162: 4122-4132. [
Medline
]
- Fallon PG, Richardson EJ, Smith
P, Dunne DW 2000a. Elevated type-1, dimished type 2 cytokines and impaired
antibody response are associated with hepatotoxicity and mortalities during
Schistosoma mansoni infection of CD-4-depleted mice. Eur J Immunol
30: 470-480.
- Fallon PG, Richardson EJ, McKenzie
GJ, McKenzie ANJ 2000b. Schistosome infection of transgenic mice defines distinct
and contrasting pathogenic roles for IL-4 and IL-13: IL-13 is a profibrotic
agent. J Immunol 164: 2585-2591. [
Medline
]
- Fallon PF, Smith P, Richardson
EJ, Jone FJ, Faulkner HC, Van Snick J, Renauld J-C, Dunne DW 2000c. Experssion
of interleukin-9 leads to Th2 cytokine-dominated responses and fatal enteropaty
in mice with chronic Schistosoma mansoni infections. Infect Immun
68: 6005-6011.
- Fanning MM, Kazura JW 1985. Further
studie on genetic variation of hepatosplenic disease and modulation in murine
schistosomiasis mansoni. Parasite Immunol 7: 213-222.
- Fanning MM, Peters PA, Davis RS,
Kazura JW, Mahmoud AAF 1981. Immunopathology of murine infection with Schistosoma
mansoni: Relationship of genetic background to hepatosplenic disease and
modulation. J Inf Dis 144: 148-153.
- Farah IO, Mola PW, Kariuki TM,
Nyindo M, Blanton RE, King CL 2000. Repeated exposure induces periportal fibrosis
in Schistosoma mansoni-infected baboons: role of TGF-b
and IL-4. J Immunol 164: 5337-5343. [
Medline
]
- Farah IO, Nyindo M, Suleman MA,
Hyaudi J, Kariuki TM, Blanton RE, Elson LH, King CL 1997. Schistosoma mansoni:
development and modulation of the granuloma after single or multiple exposures
in the baboon (Papio cynocephalus anubis). Exp Parasitol 86:
93-101. [ Medline
]
- Feldman GM, Dannenberg AM Jr.,
Seed JL 1990. Physiologic oxygen tensions limit oxidant-mediated killing of
schistosome eggs by inflammatory cells and isolated granulomas. J Leukocyte
Biol 47: 344-354.
- Ferru I, Roye O, Delacre M, Auriault
C, Wolowczuk I 1998. Infection of B-cell-deficient mice by the parasite Schistosoma
mansoni: demonstration of the participation of B cells in granuloma modulation.
Scand J Immunol 48: 233-240. [
Medline
]
- Fiore M, Moroni R, Alleva E, Aloe
L 1966. Schistosoma mansoni: influence of infection on mouse behavior.
Exp Parasitol 83: 46-54.
- Flores Villanueva PO, Harris TS,
Ricklan DE, Stadecker MJ 1994. Macrophages from schistosomal egg granulomas
induce unresponsiveness to specific cloned Th-1 lymphocytes in vitro and down-regulate
schistosomal granulomatous disease in vivo. J Immunol 152: 1847-1855.
[ Medline
]
- Flores-Villanueva PO, Zheng XX,
Strom TB, Stadecker MJ 1996. Recombinant IL-10 and IL-10/Fc treatment down-regulate
egg antigen-specific delayed hypersensitivity reactions and egg granuloma
formation in schistosomiasis. J Immunol 156: 3315-3320.
[ Medline
]
- Frizell E, Liu S-L, Abraham A,
Ozaki I, Eghball M, Sage EH, Zern MA 1995. Expression of SPARC in normal and
fibrotic livers. Hepatology 21: 847-854.
- Fugiwara M, Makino M, Watanabe
H 1988. Schistosoma mansoni: induction of severe glomerulonephritis
in female BXSB mice following chronic infection. Exp Parasitol 65:
214-221.
- Fulford AJC, Webster M, Ouma JH,
Kimani G, Dunne DW 1998. Puberty and age-related changes in susceptibility
to schistosome infection. Parasitol Today 14: 23-26.
- Gao J-L, Wynn TA, Chang Y, Lee
EJ, Broxmeyer HE, Cooper S, Tiffany HL, Westphal H, Kwon-Chung J, Murphy PM
1997. Impaired host defense, hematopoiesis, granulomatous inflammation and
type 1-type 2 cytokine balance in mice lacking CC chemokine receptor 1. J
Exp Med 185: 1959-1968. [
Medline
]
- Gaubert S, da Costa AV, Maurage
C-A, Santos Lima EC, Fontaine J, Safitte S, Minoprio P, Capron A, Grzych J-M
1999. X-linked immunodefieiency affects the outcome of Schistosoma mansoni
infection in the murine model. Parasite Immunol 21: 89-101.
- Gharib B, Abdallahi OMS, Dessien
H, Reggi MDe 1999. Development of eosinophil peroxidase activity and concomitant
alteration of the antioxidant defenses in the liver of mice infected with
Schistosoma mansoni. J Hepatol 30: 594-602. [
Medline
]
- Goes AM, Gazzinelli G, Rocha R,
Katz N, Doughty BL 1991. Granulomatous hypersensitivity to Schistosoma
mansoni egg antigens in human schistosomiasis. III. In vitro granuloma
modulation induced by immune complexes. Am J Trop Med Hyg 4:
434-443.
- Goes AM, Rezende SA, Gazzinelli
G, Doughty BL 1994. Granulomatous hypersensitivity to Schistosoma mansoni
egg antigens in human schistosomiasis. A role for prostaglandin-induced inhibition
of in vitro granuloma formation. Parasite Immunol 16:
11-18. [ Medline
]
- Green WF, Colley DG 1981. Modulation
of Schistosoma mansoni egg-induced granuloma formation: I-J restriction
of T cell mediated suppression in a chronic parasitic infection. Proc Natl
Acad Sci USA 78: 1151-1156.
- Gryseels B, de Vlas SJ 1996. Worm
burdens in schistosome infections. Parasitol Today 12: 115-119.
- Grzych J-M, Pearce E, Cheever
A, Caulada ZA, Caspar P, Heiny S, Lewis F, Sher A 1991. Egg deposition is
the major stimulus for the production of Th2 cytokines in murine schistosomiasis
mansoni. J Immunol 146: 1322-1327. [
Medline
]
- Haseeb MA, Craig JP 1997. Suppression
of the immune-response to diphtheria toxoid in murine schistosomiasis. Vaccine
15: 45-50. [
Medline
]
- Hassanein H, Hanallah S, El-Ahwany
E, Doughty B, El-Ghorab N, Badir B, Sharmy R, Zada S 2001. Immunolocalization
of intercellular adhesion molecule-1 and leukocyte functional associated antigen-1
in schistosomal soluble egg antigen-induced granulomatous hyporesponsiveness.
APMIS 109: 376-382. [
Medline
]
- Hayashi N, Matsui K, Hiroko H,
Osada Y, Mohamed RT, Nakano H, Kashiwamura S, Hyodo Y, Takeda K, Akira S,
Hada T, Higashino K, Kojima S, Nakanishi K 1999. Kupffer cells from Schistosoma
mansoni-infected mice participate in the prompt type 2 differentiation
of hepatic T cells in response to worm antigens. J Immunol 163:
6702-6711. [ Medline
]
- Henderson GS, Lu X, McCurley TL,
Colley DG 1992. In vivo molecular analysis of lymphokines involved in the
murine immune response during Schistosoma mansoni infection. II. Quantification
of IL-4 mRNA, IFN-g mRNA and IL-2 mRNA levels in
the granulomatous livers, mesenteric lymph nodes, and spleens during the course
of modulation. J Immunol 147: 2261-2269.
- Henderson GS, Nix NA, Montesano
MA, Gold D, Freeman GL, McCurley TL, Colley DG 1993. Two distinct pathologic
syndromes in male CBA/J inbred mice with chronic Schistosoma mansoni
infections. Am J Pathol 142: 703-714.
- Hernandez HJ, Stadecker MJ 1999.
Elucidation and role of critical residues of immunodominant peptide associated
with T cell-mediated parasitic disease. J Immunol 163:3877-3882.
[ Medline
]
- Hernandez HJ, Edson CM, Harn DA,
Ianelli CJ, Stadecker MJ 1998. Schistosoma mansoni: genetic restriction
and cytokine profile of the CD4+ T helper cell response to dominant epitope
peptide of major egg antigen Sm-p40. Exp Parasitol 90: 122-130.
[ Medline
]
- Hernandez HJ, Sharpe AH, Stadecker
MJ 1999. Experimental murine schistosomiasis in the absence of B7 costimulatory
molecules: reversal of elicited T cell cytokine profile and partial inhibition
of egg granuloma formation. J Immunol 162: 2884-2889.
[ Medline
]
- Hernandez HJ, Trzyna WC, Cordingley
JS, Brodeur PH, Stadecker MJ 1997c. Differential antigen recognition by T
cell populations from strains of mice developing polar forms of granulomatous
inflammation in response to eggs of Schistosoma mansoni. Eur J Immunol
27: 666-670. [
Medline
]
- Hernandez HJ, Wang Y, Stadecker
MJ 1997a. In infection with Schistosoma mansoni, B cells are required
for T helper type 2 cells responses but not for granuloma formation. J
Immunol 158: 4832-4837. [
Medline
]
- Hernandez HJ, Wang Y, Tzellas
N, Stadecker MJ 1997b. Expression of class II, but not class I, major histocompatibility
complex molecules is required for granuloma formation in infection with Schistosoma
mansoni. Eur J Immunol 27: 1170-1176. [
Medline
]
- Hesse M, Cheever AW, Jankovic
D, Wynn TA 2000. NOS-2 mediates the protective anti-inflammatory and anti-fibrotic
effects of the Th1-inducing adjuvant, IL-12, in a Th2 model of granulomatous
disease. Am J Pathol 157: 945-955. [
Medline
]
- Hesse M, Modolell M, LaFlamme
AC, Schito M, Fuentes JM, Cheever AW, Pearce EJ, Wynn TA 2001. l Differential
regulation of nitric oxide synthase-2 and arginase-1 by Type1/type2 cytokines
in vivo: granulomatous pathology is shaped by the pattern of L-arginine metabolism.
J Immunol 167: 6533-6544. [
Medline
]
- Hiatt RA, Ottesen EA, Sotomayor
ZR, Lawly TJ 1980. Serial observations of circulating immune complexes in
patients with acute schistosomiasis. J Inf Dis 142: 665-670.
- Hoffman KF, Cheever AW, Wynn TA
2000. IL-10 and the dangers of immune polariaztion. Excessive type-1 and type-2
cytokine responses induce distinct forms of lethal immunopathology in murine
schistosomiasis. J Immunol 164: 6406-6416.
- Hoffmann KF, McCarty TC, Segal
DH, Chiaramonte M, Hesse M, Davis EM, Cheever AW, Meltzer PS, Morse HC III,
Wynn TA 2001. Disease fingerprinting with cDNA microarrays reveals distinct
gene expression profiles in lethal type-1 and type-2 cytokine-mediated inflammatory
reactions. FASEB J 15: U233-U255.
- Hogan LH, Weinstock JV, Sandor
M 1999. TCR specificity in infection induced granulomas. Immunol Let
68: 115-120.
- Houba V 1979. Experimental renal
disease due to schistosomiasis. Kidney Int 16: 30-43.
[ Medline
]
- Houba V, Sturrock RF, Butterworth
AE 1977. Kidney lesions in baboons infected with Schistosoma mansoni.
Clin Exp Immunol 30: 439-449. [
Medline
]
- Hsu SYL, Hsu HF, Lust GL, Davis
JR, Eveland LK 1973. Comparative studies on the lesions caused by eggs of
Schistosoma japonicum and Schistosoma mansoni in the liver of
hamsters, guinea pigs, and albino rats. Ann Trop Med Parasitol 67:
349-356. [ Medline
]
- Hu JS, Frait KA, Reich F, Zhu
Z, Elias JA, Chensue SW 2001. IL-13 transgene state impairs mycobacterial
(Type-1) and schistosomal (Type 2) antigen-elicited responses. Cell Immunol
213: 114-121. [
Medline
]
- Jacobs W, Bogers J, Deelder A,
Marck EV 1997b. Expression of intercellular adhesion molecule-1 and lympho-cytefunction-associated
antigen-1 in experimental Schistosoma mansoni infection and in synchronous
periparticular hepatic granulomas in mice: immunohistochemistry, confocal
laser scanning microscopy, and immunoelectron microscopy. Parasitol Res
83: 405-412.
- Jacobs W, Bogers J, Deelder A,
Wéry M, Marck EV 1997a. Adult Schistosoma mansoni worms positively
modulate soluble egg antigen-induced inflammatory hepatic granuloma formation
in vivo. Am J Pathol 150: 2033-2045. [
Medline
]
- Jacobs W, Bogers JJ, Timmermans
JP, Deelder AM, Van Marck EA 1998a. Adhesion molecules in intestinal Schistosoma
mansoni infection. Parasitol Res 84: 276-280.
- Jacobs W, Deelder A, Van Marck
E 1999. Schistosomal granuloma modulation. II. Specific immunogenic carbohydrates
can modulate schistosome-egg-antigen-induced hepatic granuloma formation.
Parasitol Res 85: 14-18.
- Jacobs W, van Dam G, Bogers J,
Deelder A, Van Marck E 1998c. Schistosomal granuloma modulation. I. Schisosoma
mansoni worm antigens CAA and CCA prime egg-antigen-induced hepatic granuloma
formation. Parasitol Res 85: 7-13.
- James SL 1995. Role of nitric
oxide in parasitic infections. Microbiol Rev 59: 533-547. [
Medline
]
- Jankovic D, Sher A 1996. Initiation
and regulation of CD4+ T-cell function in host-parasite models. Chem Immunol
63: 51-65. [
Medline
]
- Jankovic D, Cheever AW, Kullberg
MC, Wynn TA, Yap G, Caspar P, Lewis FA, Clynes R, Ravetch JV, Sher A 1998.
CD4+ cell-mediated granulomatous pathology in schistosomiasis is
downregulated by a B cell-dependent mechanism requiring Fc receptor signaling.
J Exp Med 187: 619-629. [
Medline
]
- Jankovic D, Kullberg MC, Dobrowicz
D, Barbieri S, Caspar P, Paul WE, Cheever AW, Kinet J-P, Sher A 1997. FceR1-deficient
mice infected with Schistosoma mansoni display defective IL-4 production
from non-B, non-T cells as well as enhanced liver pathology. J Immunol
159: 1868-1875.
- Jankovic D, Kullberg MC, Noben-Trauth
N, Caspar P, Ward JM, Cheever AW, Paul WE, Sher A 1999. Schistosome-infected
IL-4 recepter knockout (KO) mice, in contrast to IL-4 KO mice, fail to develop
granulomatous pathology while maintaining the same lymphokine expression profile.
J Immunol 163: 337-342.
- Jankovic D, Kullberg MC, Noben-Trauth
N, Caspar P, Paul WE, Sher A 2000. Single cell analysis reveals that IL-4
receptor/Stat6 signaling is not required for the in vivo or in vitro development
of CD4+ lymphocytes with a Th2 cytokine profile. J Immunol 164:
3047-3055. [ Medline
]
- Jesus, AR, Silva A, Santant LB,
Magalhaes, A, Jesus AA, Almeida RP, Rego MA, Buratini MN, Pearce EJ, Carvalho
EM 2002. Clinical and immunologic evaluation of 31 patients with acute schistosomiasis
mansoni. J Inf Dis 185: 98-105.
- Jones JT, Kusel JR 1985. The inheritance
of responses to schistosomiasis mansoni in two pairs of inbred strains of
mice. Parasitology 90: 289-300.
- Jones JT, Breeze P, Kusel JR 1989.
Schistosome fecundity: influence of host genotype and intensity of infection.
Int J Parasitol 19: 769-777. [
Medline
]
- Jones JT, McCaffery DM, Kusel
JR 1983. The influence of the H-2 complex on responses to infection by Schistosoma
mansoni in mice. Parasitology 86: 19-30.
- Joseph AL, Boros DL 1993. Tumor
necrosis factor plays a role in Schistosoma mansoni egg-induced granulomatous
inflammation. J Immunol 151: 5461-5471. [
Medline
]
- Junqueira LCU, Bignolas G, Brentani
RR 1979. Picrosirius staining plus polarization microscopy, a specific method
for collagen detection in tissue sections. Histochem J 11: 447455.
- Kakizoe Y 1985. The influence
of Schistosoma mansoni infection on carcinogenesis of mouse livers
initiated by N-2-flurenylacetamide. Kurume Med J 32: 169-178.
[ Medline
]
- Kaplan MH, Whitfield JR, Boros
DL, Grusby MJ 1998. Th2 cells are required for the Schistosoma mansoni
egg-induced granulomatous response. J Immunol 160: 1850-1856.
[ Medline
]
- Karanja DS, Colley DG, Nahlen
BL, Ouma JH, Secor WE 1997. Evidence for immune-facilitated excretion of schistosome
eggs from patients with Schistosoma mansoni and human immunodeficiency
virus coinfection. Am J Trop Med Hyg 56: 515-521. [
Medline
]
- Kee CK, Taylor DW, Cordingley
JS, Butterworth AE, Munro AJ 1986. Genetic influence on the antibody response
to antigens of Schistosoma mansoni in chronically infected mice. Parasite
Immunol 8: 565-574. [
Medline
]
- King CL, Xianli J, Malhotra I,
Liu S, Mahmoud AAF, Oettgen HC 1997. Mice with a targeted deletion of the
IgE gene have increased worm burdens and reduced granulomatous inflammation
following primary infection with Schistosoma mansoni. J Immunol 158:
294-300. [ Medline
]
- Kloetzel K 1971. Tissue reactions
to Schistosoma mansoni ova. III. Microcirculation in the intestine
of infected mice, as studied by injection of contrast material. Rev Inst
Med Trop São Paulo 13: 51-56.
- Kloetzel K, Chieffi PP, Faleiros
J, Merluzzi TJ Filho 1977. Mortality and other parameters of concomitant infections
in albino mice: The Schistosoma-Toxoplasma model. Trop Geograph
Med 29: 407-410.
- Kloetzel K, Faleiros JJ, Mendes
SR, Stanley CT, Arias HS 1973. Concomitant infection of albino mice by Trypanosoma
cruzi and Schistosoma mansoni. Parasitological parameters. Trans
R Soc Trop Med Hyg 67: 652-658. [
Medline
]
- Knauft RF, Warren KS 1969. The
effect of calorie and protein malnutrition on both the parasite and the host
in acute murine schistosomiasis mansoni. J Inf Dis 120: 560-575.
- Knight R, Warren, KS 1973. The
interaction between Entamoeba Histolytica and Schistosoma mansoni
infections in mice. Trans R Soc Trop Med Hyg 67: 644-651.
[ Medline
]
- Kuntz RE, Malakatis GM 1955. Susceptibility
studies in schistosomiasis. II. Susceptibility of wild mammals to infection
by Schistosoma mansoni in Egypt, with emphasis on rodents. Am J
Trop Med Hyg 4: 75-89.
- La Flamme, AC, Patton EA, Pearce
EJ 2001. Role of gamma interferon in the pathogenesis of severe schistosomiasis
in Interleukin-4-deficient mice. Infect Immun 69: 7445-7452.
[ Medline
]
- Langley JG, Boros DL 1995. T-lymphocyte
responsiveness in murine schistosomiasis mansoni is dependent upon the adhesion
molecules intercellular adhesion molecule-1, lymphocyte function-associated
antigen-1 and very late antigen-4. Infect Immun 63: 3980-3986.
[ Medline
]
- Lenzi HL, Kimmel E, Schechtman
H, Pelajo-Machado M, Romanha WS, Pacheco RG, Mariano M, Lenzi JA 1998. Histoarchitecture
of schistosomal granuloma development and involution: morphogenic and biomechanical
approaches. Mem Inst Oswaldo Cruz 93 (Suppl. I):141-151.
- Lenzi HL, Kimmel E, Schechtman
H, Pelajo-Machado M, Vale BS, Panasco MS, Lenzi JA 1999. Collagen arrangement
in hepatic granuloma in mice infected with Schistosoma mansoni:dependence
on fiber radiation centers. Braz J Med Biol Res 32: 639-643.
[ Medline
] [ Lilacs
] [ SciELO
]
- Lenzi HL, Lenzi JA, Rosman FC,
PelajoMachado M, Mota EM, Panasco MS, Oliveira DN 1995. Extramedullary hematopoiesis
in murine schistosomiasis mansoni. Mem Inst Oswaldo Cruz 90:
169177.
- Lenzi HL, Lenzi JA, Sobral ACL
1987. Eosinophils favor the passage of eggs to the intestinal lumen in schistosomiasis.
Bras J Med Biol Res 20: 433-435.
- Lenzi HL, Oliveira DN, Pelajo-Machado
M, Borojevic R, Lenzi JA 1996. Coelom-associated lymphomyeloid tissue (milky
spots): site of lymphoid and myelomonocytic cell generation. Braz J Med
Biol Res 29: 19-24. [
Medline
] [ Lilacs
]
- Lenzi JA, Mota EM, Pelajo-Machado
M, Paiva RAN, Lenzi HL 1995. Calomys callosus: an alternative model
to study fibrosis in schistosomiasis mansoni. The pathology of the acute phase.
Mem Inst Oswaldo Cruz 90: 311-315.
- Lenzi JA, Pelajo-Machado M, Mota
EM, Oliveira DN, Panasco MS, Andrade ZA, Lenzi H 1998. Effects of Schistosoma
mansoni infection on Calomys Callosus coelom-associated lymphomyeloid
tissue (milky spots). Mem Inst Oswaldo Cruz 93 (Suppl. I): 13-23.
- Lenzi JA, Mota EM, Pelajo-Machado
M, Vale LS, Vela BS, Andrade ZA, Lenzi HL 2002. Intestinal fibrovascular nodules
caused by Schistosoma mansoni infection in Calomys callosus Renger,
1830 (Rodentia: cricetidae): a model of concomitant fibrosis and angiogenesis.
Mem Inst Oswaldo Cruz 97 (Suppl. I): 117-127.
- Leptak CL, McKerrow JH 1997. Schistosome
egg granulomas and hepatic expression of TNF-a
are dependent on immune priming during parasite maturation. J Immunol
158: 301-307. [
Medline
]
- Lichtenberg F von, Sadun EH, Bruce
JI 1962. Tissue responses and mechanisms of resistance in schistosomiasis
mansoni in abnormal hosts. Am J Trop Med Hyg 11: 347-356.
- Loos B 1964. Über die empfänglichkeit
deutscher kleinsäugetiere (Rodentia und Insectivora) für
experimentelle infektionen mit Schistosoma mansoni. Tropenmed Parasit
15: 55-94.
- Lopes JD, Moreira AAB, Campos
R, Kanamura HY, Hoshino-Shimzu S, Gayotto LCC, Silva, LCda 1981. Circulating
antigens, antibodies and glomerular immune complexes in mice with unisexual
Schistosoma mansoni infection. Rev Inst Med Trop São Paulo
23: 155-160. [
Medline
] [ Lilacs
]
- LoVerde PT, Amento C, Higashi
GI 1980. Parasite-parasite interaction of Salmonella typhimurium and
Schistosoma mansoni. J Inf Dis 141: 177-185.
- LoVerde PT, DeWald J, Minchella
DJ, Bosshardt WC, Damian RT 1985. Evidence for host-induced selection in Schistosoma
mansoni. J Parasitol 71: 297-301. [
Medline
]
- Lukacs NW, Boros DL 1991. Identification
of larval cross-reactive and egg-specific antigens involved in granuloma formation
in murine schistosomiasis mansoni. Infect Immunity 59: 3237-3242.
- Lukacs NW, Boros DL 1993. Lymphokine
regulation of granuloma formation in murine schistosomiasis mansoni. Clin
Immunol Immunopathol 68: 57-63. [
Medline
]
- Lukacs NW, Chensue SW, Strieter
RM, Warmington K, Kunkel SL 1994. Inflammatory granuloma formation is mediated
by TNF-a-inducible intercellular adhesion molecule-1.
J Immunol 152: 5883-5889. [
Medline
]
- Lukacs NW, Kunkel SL, Strieter
RM, Warmington K, Chensue SW 1993. The role of macrophage inflammatory protein
1a in Schistosoma mansoni egg-induced granulomatous inflammation. J
Exp Med 177: 1551-1559. [
Medline
]
- Lundy SK, Boros DL 2002. Fas ligand-expressing
B-1a lymphocytes mediate CD4+-T-cell apoptosis during schistosomal
infection: induction by interleukin 4 (IL-4) and IL-10. Infect Immun
70: 812-819. [
Medline
]
- Lundy SK, Lerman SP and Boros
DL 2001. Soluble egg antigen-stimulated T helper lymphocyte apoptosis and
evidence for cell death mediated by FasL+ T and B cells during
murine Schistosoma mansoni infection. Infect Immun 69: 271-280.
[ Medline
]
- Lwin M, Last C, Targett GAT, Doenhoff
MJ 1982. Infection of mice concurrently with Schistosoma mansoni and
rodent malarias: contrasting effects of patent S. mansoni infections
on Plasmodium chabaudi, P. yoelli and P. berghei. Ann Trop
Med Parasitol 76: 265-273. [
Medline
]
- MacDonald AS, Patton EA, La Flamme
AC, Araujo MI, Huxtable CR, Bauman B, Pearce EJ 2002. Impaired Th2 development
and increased mortality during Schistosoma mansoni infection in the
absence of CD40/CD154 interaction. J Immunol 168: 4643-4649.
[ Medline
]
- Mansy SS, Yehia HA, Hassan MM,
Hassan EA, Youssef MM, Hadi AAA, Mackenzie CD 1998. Effect of octreotide on
the pathology of hepatic schistosomiasis. Drug Res 48: 855-861.
- Maron R, Palanivel V, Weiner HL,
Harn DA 1998. Oral administration of schistosome egg antigens and insulin
b-chain generates and enhances Th2-type responses in NOD mice. Clin Immunol
Immunopathol 87: 85-92. [
Medline
]
- Marshall AJ, Rosa Brunet L, van
Gessel Y, Alcaraz A, Bliss SK, Pearce EJ, Denkers EY 1999. Toxoplasma gondii
and Schistosoma mansoni synergize to promote hepatocyte dysfunction
associated with high levels of plasma TNF-a and
early death in C57BL/6 mice. J Immunol 163: 2089-2097.
[ Medline
]
- Mathew RC, Boros DL 1986. Anti-L3T4
antibody treatment suppresses hepatic granuloma formation and abrogates antigen-induced
interleukin-2 production in Schistosoma mansoni infection. Infect
Immunity 54: 820-826.
- Mathew RC, Ragheb S, Boros DL
1990. Recombinant IL-2 therapy reverses diminished granulomatous responsiveness
in anti-L3T4-treated Schistosoma mansoni-infected mice. J Immunol
144: 4356-4361. [
Medline
]
- McClary H, Koch R, Chisari FV,
Guidotti LG 2000. Inhibition of hepatitis B virus replication during Schistosoma
mansoni infection in transgenic mice. J Exp Med 192: 289-294.
[ Medline
]
- McKenzie GJ, Fallon PG, Emson
CL, Grencis RK, McKenzie ANJ 1999. Simultaneous disruption of (IL)-4 and IL-13
defines individual roles in T helper cell Type 2-mediated responses. J
Exp Med 189: 1565-1572. [
Medline
]
- Mendlovic F, Arnon R, Tarrab-Hazdai
R, Puri J 1989b. Genetic control of immune response to a purified Schistosoma
mansoni antigen. II. Establishment and characterization of specific I-A
and I-E restricted T-cell clones. Parasite Immunol 11: 683-694.
- Mendlovic F, Tarrab-Hazdai R,
Puri DJ, Arnon R 1989a. Genetic control of immune response to a purified Schistosoma
mansoni antigen. I. Effect of MHC class II antigens on the cellular, humoral
and protective responses. Parasite Immunol 11: 667-682.
[ Medline
]
- Metwali A, Elliott D, Blum AM,
Jie L, Sabdor M, Liynch R, Noben-Trauth N, Weinstock JV 1996. The granulomatous
response in murine schistosomiasis mansoni does not switch to Th1 in IL-4-deficient
C57BL/6 mice. J Immunol 157: 4546-4533. [
Medline
]
- Metzger JM, Peterson LB 1988.
Cyclosporin A enhances the pulmonary granuloma response induced by Schistosoma
mansoni eggs. Immunopharmacol 15: 103-116. [
Medline
]
- Mola PW, Farah IO, Kariuki TM,
Nyindo M, Blanton RE, King CL 1999. Cytokine control of the granulomatous
response in Schistosoma mansoni-infected baboons: role of exposure
and treatment. Infect Immunity 67: 6565-6571.
- Montenegro SML, Miranda P, Mahanty
S, Abath FGC, Teixeira KM, Coutinho EM, Brinkman J, Gonçalves I, Domingues
LAW, Domingues ALC, Sher A, Wynn TA 1999. Cytokine production in acute versus
chronic human schistosomiasis mansoni: the cross-regulatory role of interferon-gamma
and interleukin-10 in the responses of peripheral blood mononuclear cells
and splenocytes to parasite antigens. J Inf Dis 179: 1502-1514.
- Montesano MA, Colley DG, Eloi-Santos
S, Freeman GL, Secor WE 1999a. Neonatal idiotypic exposure alters subsequent
cytokine pathology, and survival patterns in experimental Schistosoma mansoni
infections. J Exp Med 189: 637-645. [
Medline
]
- Montesano MA, Colley, DG, Freeman
GL, Secor WE 1999b. Neonatal exposure to idiotype induces Schistosoma mansoni
egg antigen-specific cellular and humoral immune responses. J Immunol
163: 898-905. [
Medline
]
- Montesano MA, Freeman GL, Gazzinelli
G, Colley DG 1990a. Expression of cross-reactive, shared idiotypes of anti-SEA
antibodies from humans and mice with schistosomiasis. J Immunol 145:
1002-1008. [ Medline
]
- Montesano MA, Freeman GL, Gazzinelli
G, Colley DG 1990b. Immune responses during human Schistosoma mansoni.
XVII. Recognition by monoclonal anti-idiotypic antibodies of several idiotypes
on a monoclonal anti-soluble schistosomal egg antigen antibody and anti-soluble
schistosomal egg antigen antibodies from patients with different clinical
forms of infection. J Immunol 145: 3095-3099. [
Medline
]
- Montesano MA, Freeman GL, Secor
WE, Colley DG 1997. Immunoregulatory idiotypes stimulate T helper 1 cytokine
responses in experimental Schistosoma mansoni infections. J Immunol
158: 3800-3804. [
Medline
]
- Morales-Montor J, Newhouse E,
Mohamed F, Baghdadi A, Damian RT 2001. Altered levels of hypothalamic-pituitary-adrenocortical
axis hormones in baboons and mice during the course of infection with Schistosoma
mansoni. J Inf Dis 183: 313-20.
- Morcos SH, Khayyal MT, Mansour
MM, Saleh S, Ishak E, Girgis NI, Dunn MA 1983. Reversal of hepatic fibrosis
after praziquantel therapy of murine schistosomiasis. Am J Trop Med Hyg
34: 314-321.
- Moreels TG, de Man JG, Rogers
JJ, de Winter, BY, Vrolix G, Herman AG, van Marck EA, Pelckmans PA 2001. Effect
of Schistosoma mansoni-induced granulomatous inflammation on murine
gastrointestinal motility. Am J Physiol Gastrointest Liver Physiol
280: G1030-G1042.
- Mountford AP, Coulson PS, Cheever
AW, Sher A, Wilson RA, Wynn TA 1999. Interleukin-12 can directly induce T-helper
1 responses in interferon-gamma (IFN-gamma) receptor-deficient
mice, but requires IFN-gamma signalling to downregulate
T-helper 2 responses. Immunolology 97: 588-594.
- Nakazawa M, Fantappie MR, Freeman
GL Jr, Eloi-Santos S, Olsen NJ, Kovacs WJ, Secor WE, Colley DJ 1997. Schistosoma
mansoni: susceptibility differences between male and female mice can be
mediated by testosterone during early infection. Exp Parasitol 815:
233-240.
- Nassauw L van, Bogers J, Van Marck
E, Timmermans JP 2001. Role of reactive nitrogen species in neuronal cell
damage during intestinal schistosomiasis. Cell Tissue Res 303:
329-336. [ Medline
]
- Natali PG, Cioli D 1976. Immune
complex nephritis in Schistosoma mansoni-infected mice. Eur J Immunol
6: 359-364. [
Medline
]
- Njenga MN, Farah, IO, Muchemi
GK, Nyindo M 1998. Peri-portal fibrosis of the liver due to natural or experimental
infection with Schistosoma mansoni occurs in the Kenyan baboon. Ann
Trop Med Parasit 92: 187-193. [
Medline
]
- Novato-Silva E, Gazzinelli G,
Colley DG 1992. Immune responses during human schistosomiasis mansoni. XVIII.
Immunologic status of pregnant women and their neonates. Scan J Immunol
35: 429-437.
- Nyindo M, Farah IO 1999. The baboon
as a non-human primate model of human schistosome infection. Parasitol
Today 15: 478-482. [
Medline
]
- Olds GR, Meneza S el, Mahmoud
AAF, Kresina TF 1989. Differential immunoregulation of granulomatous inflammation,
portal hypertension, and hepatic fibrosis in murine schistosomiasis mansoni.
J Immunol 142: 3605-3611.
- Oliveira VR, El-Cheikh MC, Aguiar
AM, Balduino A, Pinho MDB, Reis LFL, Borojevic R 2000. Schistosoma mansoni
egg-induced hepatic granulomas in mice deficient for the interferon-gamma
receptor have altered populations of macrophages, lymphocytes and connective
tissue cells. Microbes & Infect 2: 1817-1826.
- O'Regan AW, Hayden JM, Body S,
Liaw L, Mulligan N, Goetschkes M, Berman JS 2001. Abnormal pulmonary granuloma
formation in osteopontin-deficient mice. Am J Respir Crit Care Med 164:
2243-2247.
- Park MK, Hoffmann KF, Cheever
AW, Amichay D, Wynn TA, Farber JM 2001. Patterns of chemokine expression in
models of Schistosoma mansoni inflammmation and infection reveal
relationships between type 1 and type 2 responses and chemokines in vivo.
Infect Immun 69: 6755-6768.
- Patton EA, La Flamme AC, Pedras-Vasconcelos
JA, Pearce EJ 2002. Central role for Interleukin-4 in regulating nitric oxide-mediated
inhibition of t-cell proliferation and gamma interferon production in schistosomiasis.
Infect Immun 70: 177-184.
- Patton EA, Rosa Brunet L, La Flamme
AC, Pedras-Vasconcelos J, Kopf M, Pearce EJ 2001. Severe schistosomiasis in
the absence of Interleukin-4 (IL-4) is IL-12 independent. Infect Immun
69: 589-592.
- Pearce EJ, McLaren D 1983. Reappraisal
of the guinea-pig as an experimental host for studies of schistosomiasis mansoni.
Parasitology 87: 455-464.
- Pearce EJ, Cheever A, Leonard
S, Covalesky M, Fernandez-Botran R, Kohler G, Kopf M 1996. Schistosomiasis
in IL-4 deficient mice. Internat Immunol 8: 435-444.
- Pedras-Vasconcelos JA, Brunet
LR, Pearce EJ 2001. Profound effect of the absence of IL-4 on T cell responses
during infection with Schistosoma mansoni. J Leukoc Biol 70:
737-744.
- Phillips SM, Ramadan MA, Zekavat
AM, Hilliard B, Sugaya H el, Refaei M 1996. Regulation of the schistosome
granuloma and fibrosis by TGFb-1 and antifibrotic chemotherapy-safronil. Am
J Trop Med Hyg 55: 191.
- Phillips SM, Walker D, Abdel-Hafez
SK, Linette GP, Doughty BL, Perrin PJ, Fathelbab N 1987. The immune response
to Schistosoma mansoni infections in inbred rats. VI. Regulation by
T cell subpopulations. J Immunol 139: 2781-2787.
- Pinto PM, Brito CFA, Passos LKJ,
Tendler M, Simpson AJG 1997. Contrasting genomic variability between clones
from field isolates and laboratory populations of Schistosoma mansoni.
Mem Inst Oswaldo Cruz 92: 409-414.
- Portillo HA del, Damian RT 1986.
Experimental Schistosoma mansoni infection in a small new world monkey,
the saddle-back tamarin (Saguinus fusicollis). Am J Trop Med Hyg
35: 515-522.
- Powers KG, Cheever AW 1972. Comparison
of geographical strains of Schistosoma mansoni in the rhesus monkey.
Bull WHO 46: 295300.
- Prakash S, Robbins PW, Wyler DJ
1995. Cloning and analysis of murine cDNA that encodes a fibrogenic lymphokine,
fibrosin. Proc Natl Acad Sci USA 92: 2154-2158.
- Qiu B, Frait KAl, Reich F, Kornunlecki
E, Chensue, SW 2001. Chemokine expression dynamics in mycobacterial (Type-1)
and schistosomal (Type-2) antigen-elicited pulmonary granuloma formation.
Am J Pathol 155: 1503-1515.
- Rakasz E, Blum AM, Metali A, Elliot
DE, Li J, Ballas ZK, Qadir K, Lynch R, Weinstock JV 1998. Localization and
regulation of IFN-g production within the granulomas
of murine schistosomiasis in IL-4-deficient and control mice. J Immunol
160: 4994-4999.
- Rathore Q, Sacristan C, Ricklan
de, Flores Villanueva PO, Stadecker MJ 1996. In situ analysis of B7-2
costimulatory, major histocompatibility complex class II, and adhesion molecule
expression in schistosomal egg granulomas. Am J Pathol 149:
187-194.
- Remick DG, Chensue SW, Hiserodt
JC, Higashi GI, Kunkel SL 1988. Flow-cytometric evaluation of lymphocyte subpopulations
in synchronously developing Schistosoma mansoni egg and sephadex bead
pulmonary granuolomas. Am J Pathol 131: 298-307.
- Rezende SA, Lambertucci JR, Goes
AM 1997c. Role of immune complexes from patients with different clinical forms
of schistosomiasis in the modulation of in vitro granuloma reaction.
Mem Inst Oswaldo Cruz 92: 683-687.
- Rezende SA, Oliveira VR, Silva
AM, Alves JB, Goes AM, Reis LFL 1997a. Mice lacking the gamma interferon receptor
have an impaired granulomatous reaction to Schistosoma mansoni infection.
Infect Immunity 65: 3457-3461.
- Rezende SA, Silva Teixeira DN,
Drummond SC, Goes AM 1997b. IL-10 plays a role in the modulation of human
granulomatous hypersensitivity against Schistosoma mansoni eggs induced
by immune-complexes. Scand J Immunol 46: 96-102.
- Richards CS, Shade PC 1987. The
genetic variation of compatibility in Biomphalaria glabrata and Schistosoma
mansoni. J Parasitol 73: 1146-1151.
- Richter D, Harn DA, Matuschka
F-R 1995. The irradiated cercariae vaccine model: looking on the bright side
of radiation. Parasitol Today 11: 288-293.
- Ritter DM, McKerrow JH 1996.Intercellular
adhesion molecule 1 is the major adhesion molecule expressed during schistosome
granuloma formation.Infect Immun 64: 4706-4713.
- Rocha H, Oliveira VS, Oliveira
MMG de 1980. The interaction of gram negative bacteria and S. mansoni
in mice with experimental schistosomiasis. Mem Inst Oswaldo Cruz 75:
161-172.
- Rosa Brunet L, Beall M, Dunne
DW, Pearce EJ 1999b. Nitric oxide and the Th2 response combine to prevent
severe hepatic damage during Schistosoma mansoni infection. J Immunol
163: 4976-4984.
- Rosa Brunet L, Finkelman FD, Cheever
AW, Kopf MA, Pearce EJ 1997. IL-4 protects against TNF-a
mediated cachexia and death during acute schistosomiasis. J Immunol
159: 777-785.
- Rosa Brunet L, Sabin E, Cheever
AW, Kopf MA, Pearce EJ 1999a. Interleukin 5 (IL-5) is not required for expression
of a Th2 response or host resistance mechanisms during murine schistosomiasis
mansoni but does play a role in development of IL-4-producing non-T, non-B
cells. Infect Immunity 67: 3014-3018.
- Roye O, Delacre M, William IR,
Auriault C, Wolowczuk I 2001. Cutaneous interleukin-7 transgenic mice display
a propitious environment to Schistosoma mansoni infection. Parasite
Immunol 23: 133-140.
- Rumbly CA, Sugaya H, Ali Zekavat
S, Perrin PJ, Phillips SM 2001. Elimination of lymphocytes, but not eosinophils,
by FAS-mediated apoptosis in murine schistosomiasis. Am J Trop Med Hyg
65: 442-449.
- Rumbley CA, Sugaya H, Zekavat
SA, Refaei ME, Perrin PJ, Phillips SM 1999. Activated eosinophils are the
major source of Th2-associated cytokines in the schistosome granuloma. J
Immunol 162: 1003-1009.
- Rumbley CA, Zekavat AS, Sugaya
H, Perrin PJ, Ramadan MA, Phillips SM 1998. The schistosome granuloma: cha-ractrization
of lymphocyte migration, activation, and cytokine production. J Immunol
161: 4129-4137.
- Ruppel A, Rother U, Diesfeld HJ
1982. Schistosoma mansoni: development of primary infections in mice
genetically deficient or intact in the fifth component of complement. Parasitology
85: 315-323.
- Ruth JR, Warmington KS, Shang
XH, Lincoln P, Evanoff H, Kunkel SL, Chensue SW 2000. Interleukin 4 and 13
participation in mycobacterial (type-1) and schistosomal (type-2) antigen-elicited
pulmonary granuloma formation: multiparameter analysis of cellular recruitment,
chemokine expression and cytokine networks. Cytokine 12: 432-444.
- Rutitzky LI, Hernandez HJ, Stadecker
MJ 2001. Th-1-polarizing immunization with egg antigens correlates with severe
exacerbation of immunopathology and death in schistosome infection. PNAS
98: 13243-13248.
- Sabin EA, Kopf MA, Pearce EJ 1996.
Schistosoma mansoni egg-induced early IL-4 production is dependent
upon IL-5 and eosinophils. J Exp Med 184: 1871-1878.
- Sadek GA, Borges DR, Miszputen
SJ 1986. Atividade dissacaridásica intestinal da esquistossomose mansônica.
Estudo evolutivo em camundongos com diferentes cargas de infestação.
Rev Inst Med Trop São Paulo. 28: 67-73.
- Sadun EH, von Lichtenberg F, Bruce
JI 1966. Susceptibility and comparative pathology of ten species of primates
exposed to infection with Schistosoma mansoni. Am J Trop Med Hyg
15: 705-718.
- Sadun EH, von Lichtenberg F, Cheever
AW, Erickson DG 1970. Schistosoma mansoni in the chimpanzee: the
natural history of chronic infections following single and multiple exposures.
Am J Trop Med Hyg 19: 258277.
- Santos R de O, Barbosa Jr AA,
Andrade ZA 1992. Dynamics of fibrosis production and resorption in intestinal
schistosomiasis of mice. Mem Inst Oswaldo Cruz 87: 25-31.
- Schiltz JR, Olds GR, Kresina TF,
Mahmoud AAF 1988. Effect of chemotherapy on hepatic collagen and glycosaminoglycan
metabolism in Schistosoma mansoni-infected mice. Trans R Soc Trop
Med Hyg 82: 868-873.
- Schweitzer AN 1992. Alternative
patterns of MHC-restricted antibody responsiveness following intraperitoneal
immunization of inbred mice with different preparations of an 86 kilodalton
antigen of Schistosoma mansoni. Parasite Immunol 14: 267-277
- Schweitzer AN, Taylor DW 1991.
MHC-restriction of antibody responses to an 86 kilodalton antigen of Schistosoma
mansoni. Parasite Immunol 13: 261-266.
- Secor WE, Powell MR, Morgan J,
Wynn TA, Funk CD 1998. Mice deficient for 5-lipoxygenase, but not leukocyte
type 12-lipoxygenase, display alter immune responses during infection with
Schistosoma mansoni. Prostaglandins and Other Lipid Mediators
56: 291-304.
- Segal BH, Doherty TM, Wynn TA,
Cheever AW, Sher A, Holland SM 1999. The p47phox-/- mouse model
of chronic granulomatou disease has normal granuloma formation and cytokine
responses to Mycobacterium avium and Schistosoma mansoni eggs.
Infect Immun 67: 1659-1665.
- Sher A, Coffman RL, Hieny S, Scott
P, Cheever AW 1990. Interleukin 5 is required for the blood and tissue eosinophilia
but not granuloma formation induced by infection with Schistosoma mansoni.
Proc Nat Acad Sci USA 87: 61-65.
- Silva LM, Oliveira CN, Andrade
ZA 2002. Neuroschistoso-miasis. Inadequacy of the murine model. Mem Inst
Oswal-do Cruz 97: 599-600.
- Silva LCF, Borojovic R, Mourão
PAS 1990. Identification of cells responsible for synthesis of sulphated glycosaminoglycans
in schistosome-induced hepatic granulomas. Int J Exp Pathol 71:
845-856.
- Smith JH, Doughty B, Kemp WM,
Browder EJ, Mistry FD 1988. Experimental infection of the nine-banded armadillo
(Dasypus novemcinctus) with Schistosoma mansoni (Kenyan strain).
Am J Trop Med Hyg 38: 499-507.
- Souza Vidal MRF, Barbosa AA, Andrade
ZA 1993. Experimental pulmonary schistosomiasis: lack of morphological evidence
of modulation in schistosomal pulmonary granulomas. Rev Inst Med Trop São
Paulo 33: 423-429.
- Stadecker MJ, Hernandez HJ 1998.
The immune response and immunopathology in infection with Schistosoma mansoni:
a key role of major egg antigen Sm-p40. Parasite Immunol 20:
217-221.
- Stirewalt MA, Kuntz RE, Evans
AS 1951. The relative susceptibilities of the commonly-used laboratory mammals
to infection by Schistosoma mansoni. Am J Trop Med 31:
57-82.
- Takahashi S, Kobayashi M 1982.
Concanavalin A reduces liver collagen accumulation in murine schistosomiasis.
Hepato-logy 2: 249-254.
- Takahashi S, Simpser E 1981. Granuloma
collagenase and EDTA-sensitive neutral protease production in hepatic murine
schistosomiasis. Hepatology 1: 211-220.
- Tawfik AF, Carter CE, Colley DG
1986. Effects of anti-schistosomal chemotherapy on immune responses, protection
and immunity. I. Changes in cellular and humoral responses. Am J Trop Med
Hyg 35: 100-109.
- Thiongo FW, Madsen H, Ouma JH,
Andreassen J, Christensen NO 1997. Host-parasite relationships in infections
with two Kenyan isolates of Schistosoma mansoni in NMRI mice. J
Parasitol 83: 330-332.
- Todt JC, Whitfield JR, Ivard SR,
Boros DL 2000. Down-regulation of interleukin-12, Interleukin-12R expression/activity
mediates the sitch from Th1 to Th2 granuloma response during murine schistosomiasis
mansoni. Scand J Immunol 52: 385-392.
- Torrealba JF, Scorza JV, Dagert
BC 1958. The experimental infection of some wild mammals from Venezuela with
Schistosoma mansoni. Trans R Soc Trop Med Hyg 52: 565-569.
- Trzyna WC, Cordingley JS 1993.
Schistosoma mansoni: genetic non-response to p40, the major protein
antigen of the egg, reveals a novel mechanism enhancing IgM production during
infection. Parasite Immunol 15: 601-611.
- Vaillant B, Chiramonte MG, Cheever
AW, Soloway PD, Wynn TA 2001. Regulation of hepatic fibrosis and extracellular
matrix genes by the Th response: new insight into the role of tissue inhibitors
of matrix metalloproteinases. J Immunol 167: 7017-7026.
- Valadares TE, Pereira LH 1983.
Haematuria in experimental schistosomiasis mansoni. Trans R Soc Trop Med
Hyg 77: 350-353.
- Van Marck EAE, Deelder AM, Gigase
PLJ 1977. Effect of partial portal vein ligation on immune glomerular deposits
in Schistosoma mansoni-infected mice. Br J Exp Path 58:
412-417.
- Varilek GW, Weinstock JV, Williams
TH, Jew J 1991. Alteration of the intestinal innervation in mice infected
with Schistosoma mansoni. J Parasitol 77: 472-478.
- Vemgesa PB, Leese HJ 1979. Sugar
absorption by the mouse small intestine following infection with Schistosoma
mansoni. Trans R Soc Trop Med Hyg 73: 55-60.
- Verwaerde C, Thiam K, Delanoye
A, Fernandez-Gomez R, D'Halluin J-C, Auriault C 1999. Systemic delivery of
an adenovirus expressing EBV-derived vIL-10 in mice infected with Schistosoma
mansoni or Leishmania amazonensis: controversial effects on the
development of pathological parameters. Eur Cytokine Network 10:
161-170.
- Viera LQ, Moraes-Santos T 1987.
Schistosomiasis mansoni: evidence for a milder response in germ free mice.
Rev Inst Med Trop São Paulo 29: 37-42.
- Villanueva POF, Reiser H, Stadecker
MJ 1994. Regulation to T helper cell responses in experimental murine schistosomiasis
by IL-10. J Immunol 153: 5190-5199.
- Waine GJ, McManus DP 1997. Schistosomiasis
vaccine development. The current picture. BioEssays 19: 435-443.
- Wahl SM, Frazier-Jessen M, Jin
WW, Kopp JB, Sher A, Cheever AW 1997. Cytokine regulation of schistosome-induced
granuloma and fibrosis. Kidney Internat 51: 1370-1375.
- Warmington KS, Boring L, Ruth
JH, Sonstein J, Hogaboam CM, Curtis JL, Kunkel SL, Charo IR, Chensue SW 1999.
Effect of C-C chemokine receptor 2 (CCR2) knockout on type-2 (schistosomal
antigen-elicited) pulmonary granuloma formation - analysis of cellular recruitment
and cytokine responses. Am J Pathol 154: 1407-1416.
- Warren KS 1964. Experimental pulmonary
schistosomiasis. Trans R Soc Trop Med Hyg 58: 228-233.
- Warren KS 1966. The pathogenesis
of "clay-pipe stem cirrhosis" in mice with chronic schistosomiasis
mansoni, with a note on the longevity of the schistosomes. Am J Pathol
49: 477-489.
- Warren KS, Domingo EO 1970. Schistosoma
mansoni: stage specificity of granuloma formation around eggs after exposure
to irradiated cercariae, unisexual infections, or dead worms. Exp Parasitol
27: 60-66.
- Warren KS, Klein L 1969. Chronic
murine hepatosplenic schistosomiasis mansoni: relative irreversibility after
treatment. Trans R Soc Trop Med Hyg 63: 333-337.
- Warren KS, Peters PA 1967. Comparison
of penetration and maturation of Schistosoma mansoni in the hamster,
mouse, guinea pig, rabbit and rat. Am J Trop Med Hyg 16: 718-722.
- Warren KS, Simões Jr J
1966. The marmoset: a primate resistant to Schistosoma mansoni infection.
Am J Trop Med Hyg 15: 153-155.
- Warren, KS, Rosenthal MS, Domingo
EO 1969. Mouse hepatitis virus (MHV3) infection in chronic murine
schistosomiasis mansoni. Bull NY Acad Med 45: 211-224.
- Water RDe, Van Marck EAE, Fransen
JAM, Deelder AM 1988. Schistosoma mansoni: ultrastructural localization
of the circulating anodic antigen and the circulating cathodic antigen in
the mouse kidney glomerulus. Am J Trop Med Hyg 38: 87-124.
- Weinstock JV 1992. Neuropeptides
and the regulation of granulomatous inflammation. Clin Immunol Immunopathol
64: 17-22.
- Weinstock JV, Boros DL 1981. Heterogeneity
of the granulomatous response in the liver, colon, ileum, and ileal Peyer's
patches to schistosome eggs in murine schistosomiasis mansoni. J Immunol
127: 1906-1909.
- Weinstock JV, Boros DL 1983a.
Organ-dependent differences in composition and function observed in hepatic
and intestinal granulomas isolated from mice with schistosomiasis mansoni.
J Immunol 130: 418-422.
- Weinstock JV, Boros DL 1983b.
Modulation of granulomatous hypersensitivity. VI. T lymphocyte subsets influence
mast cell density in liver granulomas of Schistosoma mansoni-infected
mice. J Immunol 131: 959-961.
- Weinstock JV, Elliott D 1998.
The substance P and somatostatin interferon-gamma immunoregulatory circuit.
Neuro-immunobiology 840: 532-539.
- Weinstock JV, Blum AM, Kassab
JT 1985. Induction of granuloma modulation in murine schistosomiasis mansoni
by enteric exposure to schistosome eggs. J Immunol 135: 560-563.
- Weinstock JV, Chensue SW, Boros
DL 1983. Modulation of granulomatous hypersensitivity. V. Participation of
histamine receptor positive and negative lymphocytes in the granulomatous
response of Schistosoma mansoni-infected mice. J Immunol 130:
423-427.
- Wolowczuk I, Delacre M, Roye O,
Giannini SL, Auriault C 1997. Interleukin-7 in the skin of Schistosoma
mansoni-infected mice is associated with a decrease in interferon-g production
and leads to an aggravation of the disease. Immunology 91: 35-44.
- Wolowczuk I, Nutten S, Roye O,
Delacre M, Capron M, Murray RM, Trottein F, Auriault C 1999. Infection of
mice lacking interleukin-7 (IL-7) reveals an unexpected role for IL-7 in the
development of the parasite Schistosoma mansoni. Infect Immun
67: 4183-4190.
- Wynn TA, Cheever AW 1995. Cytokine
regulation of granuloma formation in schistosomiasis. Curr Opin Immunol
7: 505-511.
- Wynn TA, Cheever AW, Jankovic
D, Poindexter RW, Caspar P, Lewis FA, Sher A 1995a. An Il-12 based vaccination
method for preventing fibrosis induced by schistosome infection. Nature
376: 594-596.
- Wynn TA, Cheever AW, Williams
ME, Hieny S, Caspar P, Kühn R, Müller W, Sher A 1998. interleukin-10
regulates liver pathology in acute murine schistosomiasis mansoni but is not
required for immune down-modulation of chronic disease. J Immunol 160:
4473-4480.
- Wynn TA, Eltoum I, Cheever AW,
Lewis FA, Gause WC, Sher A 1993. Analysis of cytokine mRNA expression during
primary granuloma formation induced by eggs of Schistosoma mansoni.
J Immunol 151: 1430-1440.
- Wynn TA, Eltoum I, Oswald IP,
Cheever AW, Sher A 1994. Endogenous Interleukin 12 (IL-12) regulates granuloma
formation induced by eggs of Schistosoma mansoni and exogenous IL-12
both inhibits and prophylactically immunizes against egg pathology. J Exp
Med 179: 1551-1561.
- Wynn TA, Jankovic D, Hieny S,
Zioncheck K, Jardieu P, Cheever AW, Sher A 1995b. In the absence of endogenous
IFN-g, Interleukin-12 exacerbates rather than suppresses
Th2-dependent pathology. J Immunol 154: 3999-4009.
- Wynn TA, Morawetz R, Scharton-Kersten
T, Hieny S, Morse HC III, Kuhn, R, Muller W, Cheever AW, Sher A 1997. Analysis
of granuloma formation in double cytokine deficient mice reveals a central
role for IL-10 in polarizing both th1 and th2-type cytokine responses in vivo.
J Immunol 159: 5014-5023.
- Yamashita T, Boros DL 1992. IL-4
influences IL-2 production and granulomatous inflammation in murine schistosomiasis
mansoni. J Immunol 149: 3659-3664.
- Yap G, Cheever A, Caspar P, Jankovic
D, Sher A 1997. Unimpaired down-modulation of the hepatic granulomatous response
in CD8T cell and IFN-g deficient mice chronically
infected with Schistosoma mansoni. Infect Immun 65: 2583-2586.
- Zhu Y, Lukacs NW, Boros DL 1994.
Cloning of Th0 and Th2-type helper lymphocytes from liver granulomas of Schistosoma
mansoni-infected mice. Infect Immun 62: 994-999.
© 2003
Fundação Oswaldo Cruz
The following images related to this document are available:
Photo images
[oc02214t2.jpg]
[oc02214t6.jpg]
[oc02214f1-6.jpg]
[oc02214t3.jpg]
[oc02214t1.jpg]
[oc02214t5.jpg]
[oc02214t4.jpg]
|

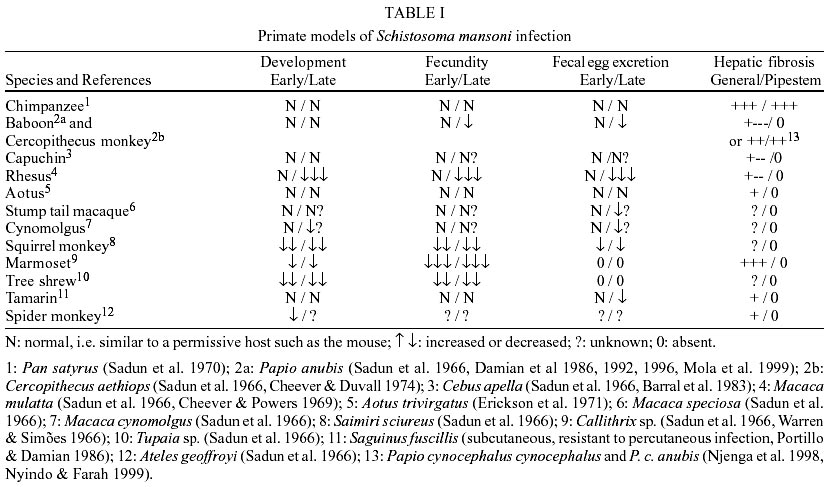{kind=link}
{kind=link}
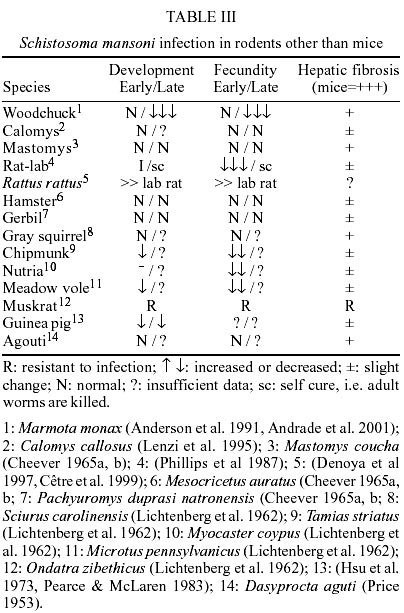{kind=link}
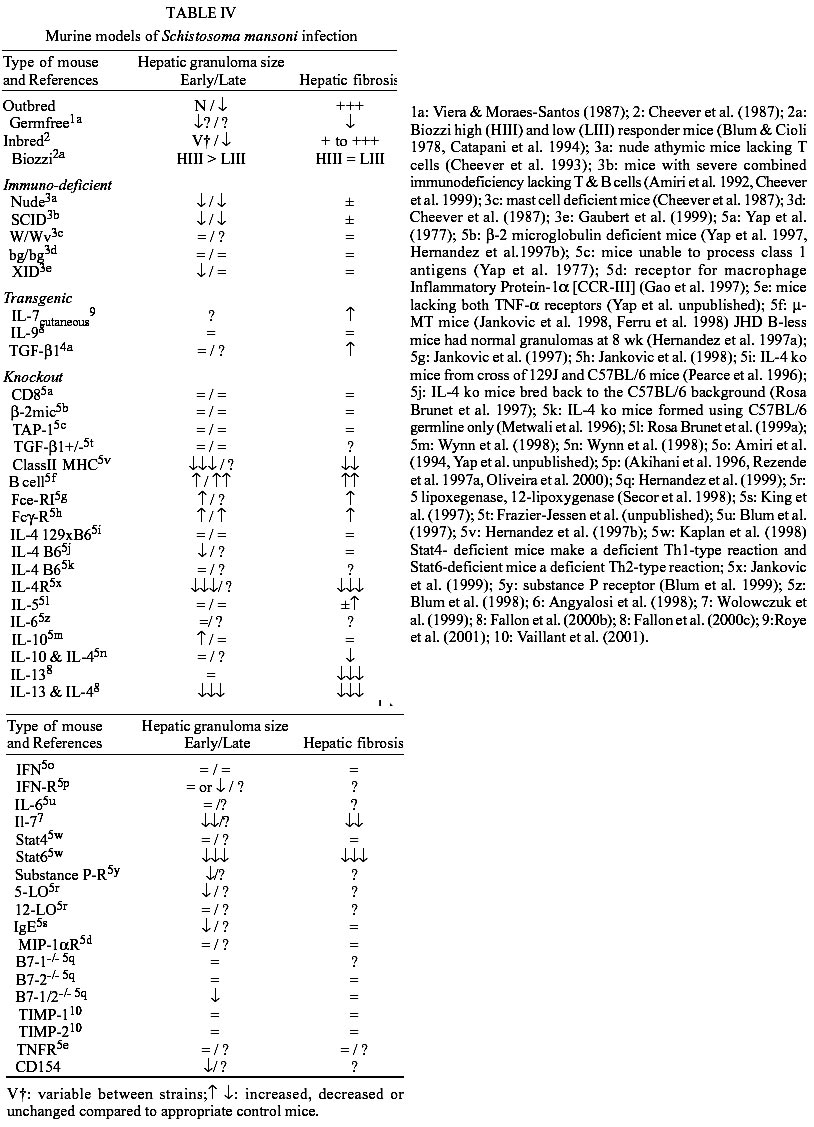{kind=link}
{kind=link}
{kind=link}
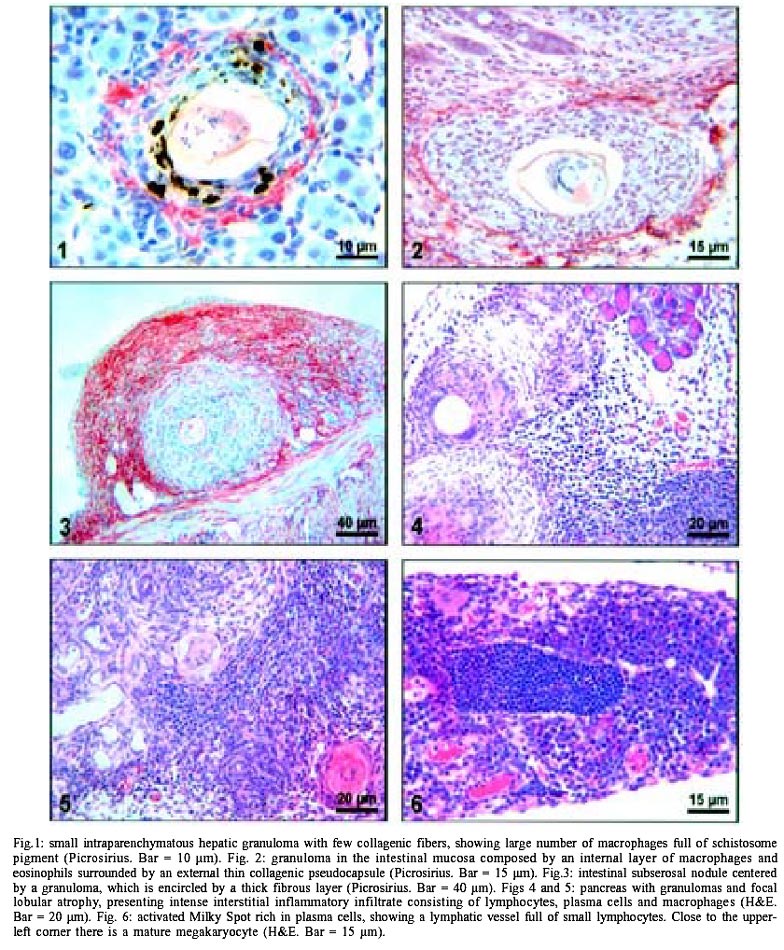{kind=link}