
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 7, 2002, pp. 941-945
|
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(7), October
2002, pp. 941-945
Enterocytozoon
bieneusi (Microsporidia) in Faecal Samples from Domestic
Animals from Galicia, Spain
B Lores+,
C del Aguila*, C Arias
Laboratorio de Parasitología,
Facultad de Ciencias, Universidad de Vigo, Vigo, España *Sección
de Parasitología, Facultad de CC Experimentales y Técnicas, Universidad
San Pablo-CEU, Madrid, España
+Corresponding author. Present address: Instituto
de Biotecnología, Universidad de Granada, Campus Fuentenueva s/n, 18071
Granada, España. Fax: +34-958-243174. E-mail: blores@ugr.es
Received 21 November 2001
Accepted 5 June 2002
This work was supported
in part by grants XUGA 30101B97 and Fundación San Pablo-CEU 06/97.
Code Number: oc02215
In this survey we examined 87
domestic animal stool samples in order to detect the possible presence of microsporidia
in animals in close contact with humans in Galicia (NW, Spain). The detection
of Enterocytozoon bieneusi spores was confirmed in faecal samples from
two dogs and one goat by polymerase chain reaction. None of the positive samples
for microsporidia in the staining method were amplified with species-specific
primers for Encephalitozoon intestinalis, E. hellem and E. cuniculi.
Four rabbits faecal samples reacted with anti-E. cuniculi serum. Our
results could indicate the importance of domestic animals as zoonotic reservoirs
of microsporidial human infections.
Key words: Enterocytozoon bieneusi
- domestic animals - faeces - zoonotic origin - Galicia - Spain
Microsporidia are protozoan parasites
belonging to the phylum Microsporidia within which exist over 1000 species classified
into approximately 100 genera. These eukaryotic obligate intracellular protozoans
have been described infecting every major animal group, especially insects,
fish, and mammals (Wittner 1999). Microsporidia have been increasingly recognized
as opportunistic pathogens of immunodeficient patients (Weber et al. 1994),
especially in Aids patients but it is also becoming increasingly common in immunocompetent
individuals (Gainzarain et al. 1998, Lores et al. 2001).
Although during the last decade numerous
data related to the epidemiology of this infection in humans and animals have
been accumulated, implying a zoonotic nature of these parasites, direct evidence
of transmission from animals to humans are still lacking (Deplazes et al. 2000).
Encephalitozoon cuniculi is
probably the most extensively studied mammalian microsporidian and has been
reported to infect a wide range of hosts, including common laboratory rodents
as well as human and non-human primates. This is the first microsporidian species
infecting humans that has been considered a zoonosis (Deplazes et al. 1996,
Didier et al. 1996) .
The first identification of E.
intestinalis in mammals other than humans was reported by Bornay et al.
(1998) in the faeces of donkeys, dogs, pigs, cow, and goat suggesting that E.
intestinalis might also be of zoonotic origin.
Enterocytozoon bieneusi is
the most frequent microsporidian found in humans, especially in Aids patients.
It has been associated mainly with chronic diarrhoea, although it has been diagnosed
in patients with other forms of immunosuppression and in immunocompetent travellers
with self-limited diarrhoea (Weber & Bryan 1994, Sobottka et al. 1995).
In addition, this pathogen has recently been detected in other natural hosts
such as pigs (Deplazes et al. 1996, Breitenmoser et al. 1999, Rinder et al.
2000), cows, goats, pigs, chickens, cats, turkeys (Bornay et al. 1998), rabbits,
dogs (del Aguila et al. 1999), and in simian immunodeficiency virus-inoculated
monkeys (Tzipori et al. 1997, Mansfield et al. 1997). Consequently, this microsporidian
infection may be more common than previously suspected.
In this study, we used microscopic,
immunologic, and molecular methods to detect microsporidial spores in faecal
samples of domestic animals from Galicia (NW, Spain). We have designed this
survey in order to expand our knowledge concerning the pathogenic role of microsporidia
in animals having close contact with humans.
MATERIALS AND METHODS
Animal stool samples - A total
of 87 faecal samples from 75 domestic animals (22 rabbits, 17 dogs, 10 cats,
4 pigs, 2 donkeys, 10 horses, 7 goats, and 3 cows) were collected during a study
conducted in rural villages in the provinces of Pontevedra and Ourense (NW,
Spain). Samples were stored in 10% formalin for several months. At the time
of this study, samples were concentrated by ethyl acetate, and stored at 4ºC
until used. Unpreserved samples were aliquoted and conserved at -80ºC for
molecular study.
Light microscopy - The samples
were stained with the Weber's chromotrope to investigate microsporidia as described
(Weber et al. 1992). A search for ova and parasites was carried out, including
ethyl acetate sedimentation, Ziehl-Neelsen stain and a specific monoclonal antibody
test for Cryptosporidium species (Merifluorâ, Meridian Diagnostics,
Inc.).
Microorganisms - E. cuniculi
(ECLD), E. hellem (CDC:V257) and E. intestinalis (CDC:V297),
were cultured on E6 monolayers to be used as controls (Visvesvara et al. 1995).
Indirect immunofluorescence test
(IIF) - Spores of E. cuniculi (ECLD), E. hellem (CDC:V257)
and E. intestinalis (CDC:V297) were harvested from culture supernatants
and processed as described (Visvesvara et al. 1991). The IIF test was performed
with specific rabbit antisera on smears from faecal and urine sediment samples.
Also, IIF was performed on control faeces known to contain E. bieneusi.
DNA extraction, purification and
PCR amplification - DNA was extracted from unpreserved faecal samples by
bead disruption of spores, followed by digestion with proteinase K, as previously
described (da Silva et al. 1999). PCR inhibitors were removed using the QIAquick
PCR Kit (Qiagen, Chatsworth, CA) according to the manufacturer's instructions.
Microsporidial SSU-rRNA coding regions
were amplified using the following species-specific primers: EBIEF1/EBIER1 for
E. bieneusi (da Silva et al. 1996), SINTF/SINTR for E. intestinalis
(da Silva et al. 1997), ECUNF/ECUNR for E. cuniculi and EHELF/EHELR for
E. hellem (Visvesvara et al. 1994). A GenAmp kit (Perkin Elmer Cetus,
Norwalk, CT, USA) was used for PCR amplification according to manufacturer's
directions. Concentration of each primer was 0.1 µg per 50 µl final
PCR reaction volume containing 1 or 0.1 µl of the purified stool sample
extract. The positive controls used included 0.003 ng per 50 µl PCR reaction
of the corresponding cloned SSU-rRNA coding region. Conditions for PCR reactions
were: denaturing at 94ºC for 30 sec in all cases, annealing at 45ºC
for 30 sec for E. intestinalis primers and at 55ºC for 30 sec for
the rest of primers, and extension at 72ºC for 90 sec. In all cases, 35
cycles were completed. Amplification products were analysed after electrophoresis
in a 2% agarose gel and visualized by staining with ethidium bromide.
Test for PCR inhibitors in purified
samples -Todetect samples with low DNA yield and to control the inhibitory
effect in the PCR reaction, purified samples were spiked with 0.003 ng
of the corresponding cloned SSU-rRNA. Amplification of a band of the correct
size indicated the removal of PCR inhibitors in the DNA purification process.
RESULTS
Light microscopy - With
Weber's chromotrope-based stain, 10 samples showed a low number of spores that
stained pinkish red with the Weber's stain and measured approximately 1-1.5
µm in length in some cases and about 2.5 µm in others. Spores observed
in two rabbit and in one donkey samples were larger and morphologically distinct
from spores found in the rest of animal stools. Clusters of microsporidia-like
spores within a vacuole inside epithelial cells were also detected in the faecal
smears of one dog and one goat. A low parasite burden determined by light microscopy
was detected in all cases, this requiring the examination of several slides
per sample.
In 10 (45.4%) rabbits samples, Eimeria
spp. was identified, coinciding with two microsporidia positive-samples. Trichuris
sp. ova were detected in two dog samples that had been found positive to microsporidia.
Trichostrongylus sp. ova were identified in one microsporidia-positive
goat sample.
Parasites found in the animal faecal
samples are summarized in the Table.
Enteric parasites were identified in most of animals studied and polyparasitism
was a generalized characteristic in all samples. Nematodes predominated over
Protozoa, and larvae and/or ova in different developmental stages were frequent
in herbivores (donkeys, horses, goats and cows).
IIF - Microsporidia spores
present in the smears from the two microsporidia-positive rabbits reacted with
the polyclonal anti-E. cuniculi serum in IIF. The few fluorescent spores
found appeared isolated and therefore difficult to diagnose.
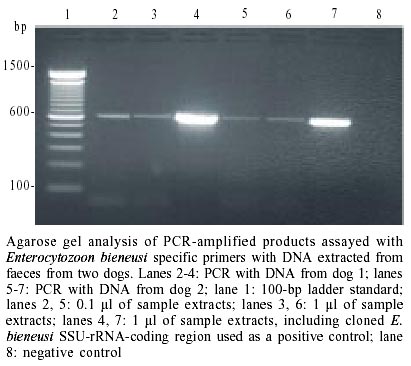
PCR amplification - PCR was
performed on unfixed faecal samples from the microsporidia-positive animals.
The DNA isolated from the five faecal samples from two dogs and one goat was
amplified with E. bieneusi specific primers and showed a diagnostic
band of 607 bp in the agarose gels (Figure).
No amplification was found in any of the five samples when species-specific
primers for E. intestinalis, E. hellem, and E. cuniculi
were used.
The donkey faecal samples that showed
Encephalitozoon-like spores did not react with any of the primers used.
To detect the possible PCR inhibitors in donkey faeces, we tested samples using
higher dilutions of template (da Silva et al. 1997) although no amplification
was observed.
DISCUSSION
The sources of microsporidia human
infections and modes of transmission, especially for E. bieneusi, remain
uncertain. Persons or animals infected with microsporidia release spores into
the environment via faecal, urine, and respiratory excretions, which all could
be possible sources of infection (Bornay et al. 1998). Although, the microsporidia
have been documented in one waterborne outbreak, the role of animals as the
cause of contamination was not elucidated. In food, surface contamination is
associated with the faecal-oral pathogens, and some data are available to indicate
that animal wastes constitute a major source of contamination (Slifko et al.
2000).
Epidemiologic data are limited, and
there are scarce data on animals as potential reservoir hosts for mi-crosporidia
that infect humans (Deplazes et al. 1996, Black et al. 1997, Mansfield et al.
1997, Bornay-Llinares et al. 1998, Didier et al. 1998).
In Spain, del Aguila et al. (1999)
were the first to report the detection of E. bieneusi in faecal
samples from wild and domestic rabbits and in domestic dogs, suggesting the
possible zoonotic role of E. bieneusi, to date considered exclusively
a human parasite.
E. bieneusi was identified
by PCR in pig faecal samples with a prevalence of 35% (Breitenmoser et al. 1999)
and, although it seems to be a common parasite in swine, no genotypes that have
been identified in humans were found. However, a recent study (Rinder et al.
2000) indicate a close relationship between E. bieneusi strains from
humans and pigs, suggesting the absence of transmission barrier between pigs
and humans for this parasite.
In the present study,using Weber's
chromotrope stain, IFI and PCR techniques, we detected microsporidian spores
in 10 faecal samples from domestic animals (2 rabbits, 2 dogs, 1 goat, and 1
donkey) from rural villages in Galicia. Microcopic examination of spores inside
the epithelial cells of faecal samples from the animals studied (dog and goat)
suggests that these animals could be hosts of E. bieneusi.
However, we do not know whether the
presence of E. bieneusi in the faecal samples are the result of an active
infection or a simple passing of the spores through the digestive tract (del
Aguila et al. 1999). Nevertheless, these results demonstrate that these animals
harboured E. bieneusi spores and shed them into the environment. This
finding will be useful to establish other possible transmission routes and to
broaden the knowledge of the human epidemiology of this infection.
To date, only E. cuniculi
infection is considered a zoonosis (Deplazes et al. 1996, Didier et al. 1996).
There are three different strains of E. cuniculi identified by Western
blot analysis of spore antigens and by random amplification of polymorphic DNA,
as well as by determination of differences in the rDNA intergenic spacer region:
strain type I includes rabbit isolates, strain type II includes murine isolates
and strain type III, dog isolates (Didier et al. 1995). The canine and rabbit
strains have been identified in three and six patients, respectively (Deplazes
et al. 1996, Didier et al. 1996, Mathis et al. 1997, Weber et al. 1997).
The above results show that in two
of the rabbits E. cuniculi spores were identified by IFI. It is unknown
whether the presence of the E. cuniculi spores released in the faeces
of these animals might signify colonization or infection by the parasite, or
simply passage through the digestive tract. This could also be a disseminated
infection and thus the spores appeared in the faeces by urine contamination.
In the faeces of a donkey, we found
spores with a size compatible with the microsporidia of the genus Encephalitozoon,
but these had no amplification by PCR of the DNA studied. The application of
the PCR technique in the analysis of faecal samples should be performed with
caution to avoid false negatives. If the parasite-DNA concentration is low,
the amplification requires large volumes of sample in the reaction. Nevertheless,
the presence of certain samples of PCR inhibitors allows adequate amplification
only when these are more diluted (da Silva et al. 1997, Ommbruck et al. 1997).
In this case, despite the repeated reactions with different DNA dilutions, amplification
does not occur with the species-specific primers assayed.
Furthermore, the high degree of parasitism
found should be emphasized, these being multiple infections mostly by nematode
parasites. Microsporidians are intracellular parasites ranked among the protists,
which means that they are eukaryotic and unicellular. They exhibit a number
of important, unique features which are the basis for the systematic ranking
of microsporidia, which are knowledged to constitute a phylum of their own (Wittner
1999). In particular, in the area where this study was made, helminthosis predominates
over protozoosis, precisely the opposite of the situation found in other surveys
in Spain, and consistent with the research performed on the presence of intestinal
parasites in diverse human populations and in other mammals in Galicia (Arias
1980). The area studied (NW, Spain), with a sub-humid Mediterranean weather-type
with an Atlantic tendency (Carballeira et al. 1983), includes an environment
favourable for the development of infective stages of different nematode species.
The presence of microsporidia in
the natural infections of animals are important for the study of clinically
significant disease in humans (Rabeneck et al. 1995). With the recent improvements
in diagnostic methods for detecting microsporidia, reports are being published
about this infection, and as more is learned about the epidemiology, immunology,
and pathology of microsporidiosis, advances in the prevention and control of
microsporidiosis are more likely in susceptible mammalian hosts (Didier et al.
2000).
The detection of new Enterocytozoon
genotypes in faecal samples of domestic animals, together with recent reports
of detection of E. bieneusi in environmental samples, suggests that microsporidia
of the genus Enterocytozoon is ubiquitous and has many genotypes in various
infected animal species (Mathis et al. 1999).
In zoonotic studies, it is important
to investigate all the possible reservoirs for each causal pathogenic agent
in any geographical area (Lores et al. 1994) and thus it is crucial to confirm
the zoonotic role of E. bieneusi, as this fact may change the established
idea of E. bieneusi as a human parasite and possibly characterize it
as a zoonotic parasite. As microsporidia are released into the environment via
stool, urine and respiratory excretion, possible sources of infection may be
persons or animals infected with this group of parasites (del Aguila et al.
1999).
Finally, the role of animals in the
transmission of microsporidial human infections requires further study. Although
animals have been implicated in some opportunistic infections among HIV-infected
persons, the overall risk of transmission from contact with domestic animals
is unknown, but may be further reduced. Nevertheless immunodeficient patients,
especially with Aids, should be informed with practical suggestions designed
to reduce this low risk.
ACKNOWLEDGMENTS
To Florentina Martinez and Adel Vázquez
for help in collecting samples. To Dr A Osuna, Instituto de Biotecnología,
Universidad de Granada, for helpful discussion.
REFERENCES
- Arias Fernández C 1980.
Parasitismo en Galicia y Tratamiento Humano y Experimental de la Trichurosis,
Thesis, Universidad de Santiago de Compostela, Spain, 383 pp.
- Black SS, Steinohrt LA, Bertucci
DC, Rogers LB, Didier ES 1997. Encephalitozoon hellem in budgerigars
(Melopsittacus undulatus). Vet Pathol 34: 189-198.
[ Medline
]
- Bornay-Llinares F, da Silva A,
Moura H, Schwartz D, Visvesvara G, Pieniazek N, Cruz-López A, Hernández-Jaúregui
P, Guerrero J, Enriquez F 1998. Immunologic, microscopic, and molecular evidence
of Encephalitozoon intestinalis (Septata intestinalis) infection
in mammals other than humans. J Infect Dis 178: 820-826.
[ Medline
]
- Breitenmoser AC, Mathis A, Burgi
E, Weber R, Deplazes P 1999. High prevalence of Enterocytozoon bieneusi
in swine with four genotypes that differ from those identified in humans.
Parasitology 118: 447-453.
- Canning E, Lom J 1986. The
Microsporidia of Vertebrates, Academic Press, Inc., New York, 289 pp.
- Carballeira A, Devesa C, Returto
R., Santillán F, Ucieda F 1983. Bioclimatología de Galicia,
Fundación Barrie de la Maza, La Coruña, Spain.
- Da Silva AJ, Schwartz DA, Visvesvara
GS, Moura H, Slemenda SB, Pieniazek NJ 1996. Sensitive PCR diagnosis of infection
by Enterocytozoon bieneusi (microsporidia) using primers based on the
region coding for small-subunit rRNA. J Clin Microbiol 34: 986-987.
- Da Silva AJ, Slemenda SB, Visvesvara
GS, Schwartz DA, Willcox CM, Wallace S, Pieniazek NJ 1997. Detection of Septata
intestinalis (Microsporidia) infections using polymerase chain reaction
primers targeting the small subunit ribosomal RNA codin region. Mol Diag
2: 47-52.
- Da Silva AJ, Bornay-Llinares FJ,
Moura H, Slemenda S, Turtle J, Pieniazek NJ 1999. Fast and reliable extraction
of protozoan parasite DNA from fecal specimens. Mol Diag 4: 57-64.
- Del Aguila C, Izquierdo F, Navajas
R, Pieniazek NJ, Miró G, Alonso A, da Silva A, Fenoy S 1999.Enterocytozoon
bieneusiin animals: rabbits and dogs as new hosts. J Eukaryot Microbiol
46: 8-9S.
- Deplazes P, Mathis A, Muller C,
Weber R 1996. Molecular epidemiology of Encephalitozoon cuniculi and
first detection of Enterocytozoon bieneusi in fecal samples of pigs.
J Eukaryot Microbiol 43: S93.
- Deplazes P, Mathis A, Weber R
2000. Epidemiology and zoonotic aspects of microsporidia of mammals and birds.
Contrib Microbiol 6: 236-260.
- Didier E, Didier P, Snowden K,
Shadduck J 2000. Micros-poridiosis in mammals. Microbes and Infection
2: 709-720.
- Didier ES, Snowden K, Shadduck
J 1998. The biology of microsporidian species infecting mammals. Adv Parasitol
40: 279-316.
- Didier ES, Visvesvara GS, Baker
MD, Rogers LB, Bertucci DC, de Groote MA, Vossbrinck CR 1996. A microsporidian
isolated from an AIDS patient corresponds to Encephalitozoon cuniculi
III, originally isolated from domestic dogs. J Clin Microbiol 34:
2835-2837. [ Medline
]
- Didier ES, Vossbrick CR, Baker
MD, Rogers LB, Bertucci DC, Shadduck JA 1995. Identification and characterization
of three Encephalitozoon cuniculi strains. Parasitology 111:
411-421.
- Gainzarain C, Canut A, Lozano
M, Labora A, Carreras F, Fenoy S, Navajas R, Pieniazek NJ, da Silva AJ, del
Aguila C 1998. Detection of Enterocytozoon bieneusi in two human immunodeficiency
virus-negative patients with chronic diarrhea by polymerase chain reaction
in duodenal biopsy specimens and review. Clin Infect Dis 27: 394-398.
[ Medline
]
- Lores B, Arias C, Lòpez-Miragaya
I, Torres J, Fenoy S, del Aguila C 2001. Molecular diagnosis of intestinal
microsporidiosis in pediatric patients from Vigo (NW, Spain). Res Rev Parasitol
61: 43-49.
- Lores B, Pascual S, Arias C 1994.
Parasites and Zoos. Parasitol Today 10: 357.
- Mansfield KJ, Carville A, Hebert
D, Chalifoux L, Shvetz D, Lin KC, Tzipori S, Lackner A 1997. Identification
of an Enterocytozoon bieneusi-like microsporidian in simian-immunodeficiency-virus-inoculated
macaques with hepato-biliary disease. Am J Pathol 150: 1395-1405.
[ Medline
]
- Mathis A, Breitenmoser AC, Deplazes
P 1999. Detection of new Enterocytozoon genotypes in faecal samples
of farm dogs and a cat. Parasite 6: 189-193. [
Medline
]
- Mathis A, Michel M, Kuster H,
Muller C, Weber R, Deplazes P 1997. Two Encephalitozoon cuniculi strains
of human origin are infectious to rabbits. Parasitology 114:
29-35.
- Ombrouck C, Ciceron L, Biligui
S, Brown S, Marechal P, Van Gool T, Datry A, Danis M, Desportes-Livage I 1997.
Specific PCR assay for direct detection of intestinal microsporidia Enterocytozoon
bieneusi and Encephalito-zoon intestinalis in fecal specimens from
human immunodefi-ciency virus-infected patients. J Clin Microbiol 35:
652-655. [ Medline
]
- Rabeneck L, Genta RM, Gyorkey
F, Clarridge JE, Gyorkey P, Foote LW 1995. Observations on the pathological
spectrum and clinical course of microsporidiosis in men infected with the
human immunodeficiency virus: follow-up study. Clin Infect Dis 20:
1229-1235. [ Medline
]
- Rinder H, Thomschke A, Dengjel
B, Gothe R, Loscher T, Zahler M 2000. Close genotypic relationship between
Entero-cytozoon bieneusi from humans and pigs and first detection in
cattle. J Parasitol 86: 185-188.
- Slifko T, Smith HV, Rose JB. 2000.
Emerging parasite zoonoses associated with water and food. Int J Parasitol
30: 1379-1393.
- Sobottka I, Albrecht H, Schottelius
J, Schmetz C, Bentfeld M, Lauff R, Schwartz DA 1995. Self limited travelers
diarrhea due to a dual infection with Enterocytozoon bieneusi and Cryptosporidium
parvum in an immunocompetent HIV-negative child. Eur J Clin Microbiol
Infect Dis 14: 919-920.
- Tzipori S, Carville A, Widmer
G, Kotler D, Mansfield K, Lackner A 1997. Transmission and establishment of
persistent infection of Enterocytozoon bieneusi, derived from a human
with AIDS, in simian immunodeficiency virus-infected rhesus monkeys. J
Infect Dis 175: 1016-1020. [
Medline
]
- Visvesvara GS, Leitch GJ, Moura
H, Wallace S, Weber R, Bryan RT 1991. Culture, electron microscopy, and immunoblot
studies on a microsporidian parasite isolated from the urine of a patient
with AIDS. J Protozool 38: 105S-111S. [
Medline
]
- Visvesvara GS, Leitch GJ, da Silva
AJ, Croppo GP, Moura H, Wallace S, Slemenda SB, Schwartz DA, Moss D, Bryan
RT, Pieniazeck NJ 1994. Polyclonal and monoclonal antibody and PCR-amplified
small-subunit rRNA identification of a microsporidian, Encephalitozoon
hellem, isolated from an AIDS patient with disseminated infection. J
Clin Microbiol 32: 2760-2768. [
Medline
]
- Visvesvara GS, da Silva AJ, Croppo
GP, Pieniazek NJ, Leitch GJ, Ferguson D, de Moura H, Wallace S, Slemnda S,
Tyrell I, Moore DF, Meador J 1995. In vitro culture and serologic and molecular
identification of Septata intestinalis isolated from urine of a patient
with AIDS. J Clin Microbiol 33: 930-936. [
Medline
]
- Weber R, Bryan RT 1994. Microsporidial
infections in immunodeficient and immunocompetent patients. Clin Infect
Dis 19: 517-521.
- Weber R, Bryan RT, Owen RL, Willcox
CM, Gorelkin L, Visvesvara GS 1992. Improved light-microscopical detection
of Microsporidia spores in stool and duodenal aspirates. N Engl J Med 326:
161-166. [ Medline
]
- Weber R, Bryan RT, Schwartz DA,
Owen RL 1994. Human microsporidial infections. Clin Microbiol Rev 7:
426-461. [ Medline
]
- Weber R, Deplazes P, Flepp M,
Mathis A, Baumann R, Sauer B, Kuster H, Luthy R 1997. Cerebral microsporidiosis
due to Encephalitozoon cuniculi in a patient with human immunodeficiency
virus infection. New Engl J Med 336: 474-478.
- Wittner M 1999. The Microsporidia
and Microsporidiosis, ASM Press, Washington, D.C., 553 pp.
© 2003
Fundação Oswaldo Cruz
The following images related to this document are available:
Photo images
[oc02215t1.jpg]
[oc02215f1.jpg]
|

{kind=link}
{kind=link}