
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 7, 2002, pp. 971-977
|
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(7), October
2002, pp. 971-977
Field Trials of
an Improved Cost-effective Device for Detecting Peridomestic Populations of
Triatoma infestans (Hemiptera: Reduviidae) in Rural Argentina
GM Vazquez-Prokopec, LA Ceballos,
OD Salomón*, RE Gürtler/+
Laboratorio de Ecología General,
Departmento de Ecología, Genética y Evolución, Universidad
de Buenos Aires, Ciudad Universitaria, C1428EHA Buenos Aires, Argentina *Centro
Nacional de Diagnóstico e Investigación en Endemo-epidemias, Administración
Nacional de Laboratorios e Institutos de Salud "Dr. Carlos G. Malbrán",
Buenos Aires, Argentina
+Corresponding author. Fax: +54-11-4576.3384.
E-mail: gurtler@bg.fcen.uba.ar
This study was
supported by UNDP/World Bank/WHO (TDR) from the Special Program for Research
and Training in Tropical Diseases, and the University of Buenos Aires.
Received 12 April 2002
Accepted 12 July 2002
Code Number: oc02220
An improved device for detecting
peridomestic Triatoma infestans consisting of one-liter recycled Tetra
Brik milk boxes with a central structure was tested using a matched-pair study
design in two rural areas in Argentina. In Olta (La Rioja), the boxes were installed
beneath the thatched roofs and on the vertical wooden posts of each peridomestic
structure. After a 5-month exposure, at least one of the recovered boxes detected
88% of the 24 T. infestans-positive sites, and 86% of the 7 negative
sites by timed manual collections at baseline. In Amamá (Santiago del
Estero), the boxes were paired with the best performing prototype tested before
(shelter unit). After 3 months, some evidence of infestation was detected in
89% (boxes) and 79% (shelters) of 18-19 sites positive by timed collections,
whereas 19% and 16% of 32 negative sites were positive, respectively. Neither
device differed significantly in the qualitative or quantitative collection
of every sign of infestation. The installation site did not modify significantly
the boxes' sampling efficiency in both study areas. As the total cost of each
box was half as expensive as each shelter unit, the boxes are thus the most
cost-effective and easy-to-use tool for detecting peridomestic T. infestans
currently available.
Key words: Triatoma infestans
- Chagas disease - surveillance - sampling - vector control - Argentina
Triatoma infestans (Klug),
the main vector of Chagas disease in Argentina, Bolivia, Brazil, Paraguay, Peru
and Uruguay, is the target of an elimination program called The Southern Cone
Initiative (Schmunis et al. 1996). T. infestans infests domestic and
peridomestic habitats almost exclusively (Zeledón & Ravinovich 1981,
Dias 1991), with sylvatic foci apparently existing only in Bolivia (Dujardin
et al. 1987, Noireau et al. 2000). Although the transmission of Trypanosoma
cruzi (Kinetoplastida: Tripanosomatidae) to humans mostly occurs in human
habitations (the domestic environment)(Cecere et al. 1999, Cohen & Gürtler
2001), peridomestic foci of T. infestans are very frequent and the most
likely source of the bugs that reinfest human habitations after insecticide
spraying (Dujardin et al. 1996, Cecere et al. 1997). Early detection of peridomestic
sites reinfested by triatomine bugs is thus essential to assess the effects
of control actions and to establish the need for additional operations.
The standard method to assess the
occurrence and intensity of infestation by triatomine bugs in domestic and peridomestic
sites has been timed manual collections using an irritant spray (the "flushing-out"
method) (Schofield 1978, Rabinovich et al. 1995). However, this method is costly,
requires skilled staff, and lacks sensitivity and precision. An artificial shelter
unit proved to be more sensitive in the detection of peridomestic foci of T.
infestans than segments of bamboo cane lined with pleated paper or timed
manual collections with an irritant (De Marco et al. 1999, Gürtler et al.
2001b). The shelter units accounted for several features of triatomine behavior
(negative phototaxis, thigmotaxis, and preference for dry sites) and were resistant
to weather conditions. However, they were expensive for large-scale triatomine
surveillance programs, and because of the rigid plastic structure, the shelter
units were not easily adaptable to many pe-ridomestic sites differing in physical
structure. Our general aim has been to improve the cost-effectiveness of the
sensitive shelter units by using recycled, appropriate materials that could
be easily handled by the affected rural populations. Such a tool is also indispensable
to study the spatial and temporal dynamics of reinfestation at a community-wide
scale.
In this study, we describe a simpler,
equally sensitive and less expensive device than the shelter unit for detecting
peridomestic triatomine foci. In two different rural areas in Argentina, we
conducted matched-paired trials to assess the devices' effectiveness at peridomestic
sites determined to be positive and negative for T. infestans by timed
manual collections, and whether the installation site modified their sensitivity.
MATERIALS AND METHODS
Devices - The new devices
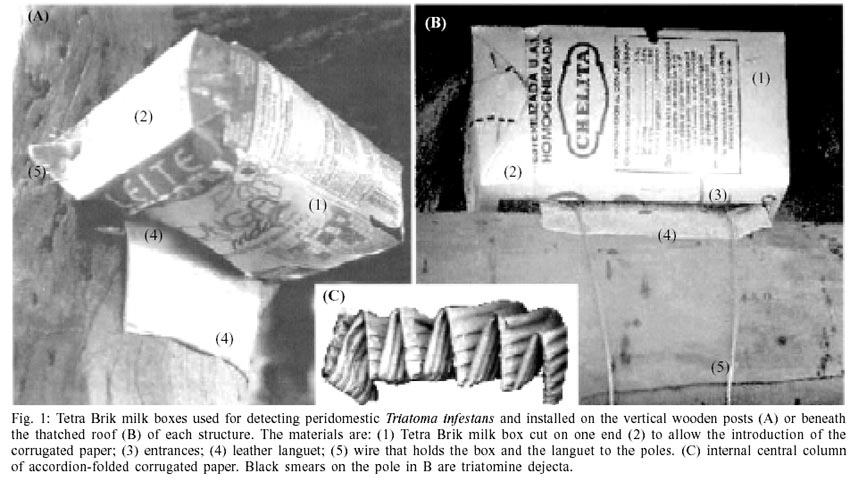
were one-liter, recycled Tetra BrikÔ (Tetra
Pak Corp.) milk boxes measuring 16 x 9.5 x 6 cm (length, width, depth), hereafter
called `box'. Two versions of these boxes, which differed only in the location
of the entrances, were designed for installation beneath the beams supporting
the thatched roof, or on the vertical wooden posts of each structure (Fig.
1). Internally, the boxes contained a central column (20 cm long and 4.5
cm in section) of accordion-folded corrugated paper (Fig.
1C), and a piece of corrugated paper lining the internal walls. All boxes
had been washed with diluted detergent before field use, whereas other component
materials had not been used or treated with any bug insecticide or attractant.
To install the boxes on the vertical
posts, a single strand of wire was passed through each of two holes made on
the two folded triangles located at the base and top of the box (Fig.
1A). The wire also held in place a 10 x 12 cm piece of commercially available
bovine leather (the languet), with the rough surface facing upwards and covering
the 10 x 1 cm entrance. The entrance was located at the bottom to intercept
the bugs when they returned to the refuges after feeding or host-seeking activities
(Fig. 1A). When the box was placed
beneath and parallel to the horizontal beams supporting the thatched roof (Fig.
1B), the two entrances on the sides were also coated with a 10 x 12 cm piece
of leather bounded flush to the beams with wire.
De Marco et al. (1999) described
the shelter units. They consisted of a black plastic, wide-mouthed jar (19 cm
high and 10 cm diameter) with a screw cap at the top and two openings at the
bottom, from which a languet of leather coating the floor extended to the outside.
Internally, the shelter units contained a removable central column (Fig.
1C) and the walls were lined with corrugated paper.
Study sites -As part of a
larger research project on the ecology and control of T. infestans, we
first conducted a trial of the boxes in rural houses around Olta (30.3ºS,
66.2ºW), La Rioja, Argentina, from late April 1999 to November 1999. The
area belongs to the Monte province of the phytogeographic Chaco region, and
is semi-arid with a thorn forest (Cabrera & Willink 1980). Olta is 471 m
above sea level, and has a mean annual temperature of 19.3ºC, and the average
annual rainfall is about 450 mm. The houses had been sprayed with beta-cyfluthrin
approximately 5-6 years before this trial. In view of the positive results obtained
in Olta, we conducted a matched-pair field trial of boxes and shelter units
in Amamá and nearby villages (27.1oS, 63oW), Santiago
del Estero, Argentina, between December 1999 and March 2000. The area belongs
to the Chaco province of the phytogeographic Chaco region (Cabrera & Willink
1980). Its environmental characteristics and history of infestation by T.
infestans were described previously (Gürtler et al. 1999).
Study design - In Olta, two
skilled bug collectors from the National Vector Control Program (NVCP) assessed
the intensity of peridomestic infestation by T. infestans and other triatomine
species from 20 April to 6 May 1999. The search was made using an irritant agent
(0.2% tetramethrin, Icona, Argentina) for 30 min per house, as described by
Gürtler et al. (1995). All triatomine bugs were removed, identified to
species and counted by stage, as described by Canale et al. (2000). Another
person placed one or two pairs of numbered boxes per site in late June 1999.
In total, 39 pairs of boxes were installed beneath the roofs and on the posts
in 34 peridomestic sites at 26 houses. The peridomestic sites included 23 goat
corrals and 2 chicken coops in which T. infestans had just been captured
by timed manual collections, and 9 apparently uninfested sites (4 goat corrals,
1 pig pen, and 4 chicken coops).
All boxes were inspected for evidence
of infestation following standard procedures between 14 and 18 November 1999
(Gürtler et al. 2001b). Briefly, the boxes were handled inside a deep plastic
tray to minimize the loss of eggs or other signs, and the number of triatomine
bugs, exuviae, eggs and dejecta in the interior and exterior of each device
were counted and recorded. Triatomine-like dejecta from the boxes were tested
by the phenolphthalin (Kastle-Meyer) test (Gürtler et al. 2001a) to confirm
the presence of heme (which is absent from other non-hematophagous arthropods'
feces). A maximum of 10 dejecta per device was tested to verify that at least
one was phenolphthalin-positive.
In Amamá, one or two matched
pairs of shelter units and boxes were installed between 10 and 16 December 1999.
A total of 79 pairs (37 beneath the roofs and 42 on the posts) were installed
in 56 peridomestic sites at 40 houses. The 24 infested sites included 6 goat
corrals, 4 pig pens, 9 storerooms, 1 oven, and 4 chicken coops in which T.
infestans had been collected by timed manual collections in April 1999.
The 32 negative sites included 9 goat or sheep corrals, 12 pig pens, 2 storerooms,
2 ovens, 6 chicken coops, and 1 pile of wood. Whenever possible, a pair of shelter
units and boxes was installed beneath the roof and another pair on the posts,
but in sites that did not allow a double installation, the pair that adapted
best was chosen. Between 14 and 27 March 2000, two skilled bug collectors from
NVCP searched for triatomines in all bedrooms and peridomestic areas of 114
houses using 0.2% tetramethrin (Icona, Argentina) for 30 min per house. All
triatomine bugs were processed as in Olta. Simultaneously with the timed manual
collections, all boxes and shelter units were inspected for evidence of triatomine
infestation by another team. For timed manual collections, the term "infested"
or "positive" meant finding at least one live or moribund T. infestans.
For the devices, these terms meant that at least one sign of infestation (i.e.,
T. infestans bugs, eggs or exuvie, or triatomine fecal smears) was detected.
Data analysis - Given the
matched-pairs study design, each pair of devices was taken as the sampling unit
to test their relative effectiveness. Pairs with one missing device were excluded
from analysis. In the absence of prior evidence, the sensitivity of devices
was tested by the twotailed binomial test (Zar 1996). The recovery rate of each
device was tested by the McNemar test for paired data (Zar 1996). In Olta, the
sampling efficiency of the boxes according to installation site was measured
by the log-transformed ratios of the numbers of bugs or each sign of infestation
recovered from each box placed on the post (x) to those in the matched box beneath
the roof (y) of each peridomestic site {log10[(x + 1)/(y + 1)]}.
The antilog of the mean log ratio is the geometric mean ratio. Using a similar
procedure for data collected in Amamá, the relative sampling efficiency
of the devices was measured by the log-transformed ratios of the numbers of
bugs or each sign of infestation recovered from each box (x) to those in the
matched shelter unit (y) at each installation site {log10[(x + 1)/(y
+ 1)]}. Variations in the density of triatomine dejecta among component materials
were studied by Kruskal-Wallis test (Zar 1996).
RESULTS
In Olta, the total catch of T.
infestans by timed manual collections from 25 infested peridomestic sites
was 199 bugs (median, 3 bugs per site; first and third quartiles, Q1-Q3, 2-9.3)
in late April 1999. In November 1999, the boxes that had been installed beneath
the roofs were recovered significantly more frequently than those on the vertical
posts (90% to 69%, respectively; McNemar c2=
6.13; degrees of freedom, df = 1, P = 0.013). All of the missing
devices had been placed in goat or sheep corrals.
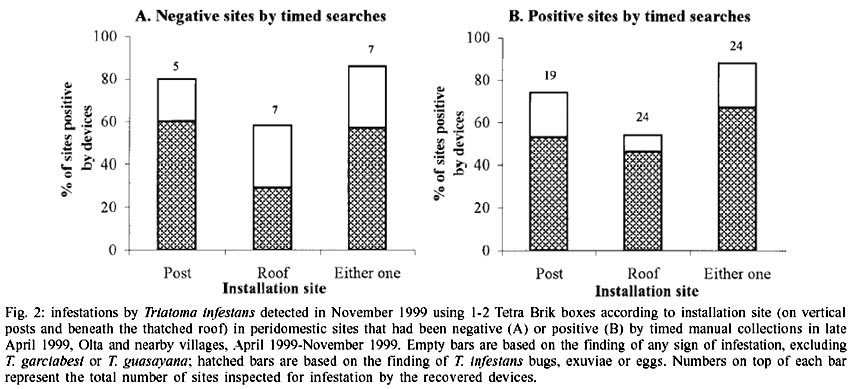
At least one of the recovered boxes
detected some evidence of triatomine infestation in 88% of the positive sites,
and in 86% of the sites that had been negative by timed collections in late
April 1999 (Fig. 2). Both in positive
and negative sites, the boxes on the posts were qualitatively more sensitive
in detecting T. infestans bugs or any sign of infestation than those
beneath the thatched roofs, although not significantly so (two-tailed binomial
tests, P > 0.2).
The frequency distribution of every
sign in the positive devices was significantly different from a random distribution:
T. infestans bugs (c2 = 58.5, df
= 16, P < 0.005), exuviae (c2=
27.3, df = 9, P < 0.005), eggs (c2
= 296.6, df = 19, P < 0.005) or triatomine dejecta (c2
= 240.0, df = 30, P < 0.005); the variance to mean ratios (11,
2, 18, and 10, respectively) indicated strong aggregation. The boxes collected
a similar total number of signs of infestation when placed beneath the roofs
(443) or on the posts (393). When placed beneath the roofs rather than on the
posts, the boxes' sampling efficiency did not differ significantly for T.
infestans bugs (mean log ratio ± 95% confidence limits (C.L.), 0.03
± 0.29), exuviae (0.18 ± 0.43), eggs (0.09 ± 0.39) or triatomine
dejecta (0.17 ± 0.25). The 67 bugs collected from the boxes in five negative
sites were 76% I-II instar nymphs, 3% III-IV, 9% V, and 12% adults, whereas
in the positive sites the stage structure of the 78 bugs collected was 49% I-II,
13% III-IV, 17% V, and 21% adults. Both stage structures were significantly
different (c2 = 32.6; df
= 3; P < 0.001). The density of triatomine dejecta per 100 cm2
in the languet (median, 1.6; Q1-Q3, 0.8-2.5) was significantly larger by at
least one order of magnitude than in the accordion (median, 0.4; Q1-Q3, 0.2-0.8),
wall lining (median, 0.4; Q1-Q3, 0.2-1.1) and the external surface (median,
0.6; Q1-Q3, 0.2-1.5)(Kruskal-Wallis, c2 =
10.1; df = 3; P = 0.018).
In Amamá, the total timed
manual catch of T. infestans from 24 infested peridomestic sites was
194 bugs in March 2000 (median per site, 5 bugs; Q1-Q3, 2-14). There were no
significant differences in the recovery rate of the 79 pairs of boxes and shelter
units when placed either beneath the thatched roof (84% and 92%, respectively;
McNemar c2= 0.57; df = 1; P
= 0.45), or on the vertical posts (93% and 88%, respectively; McNemar c2=
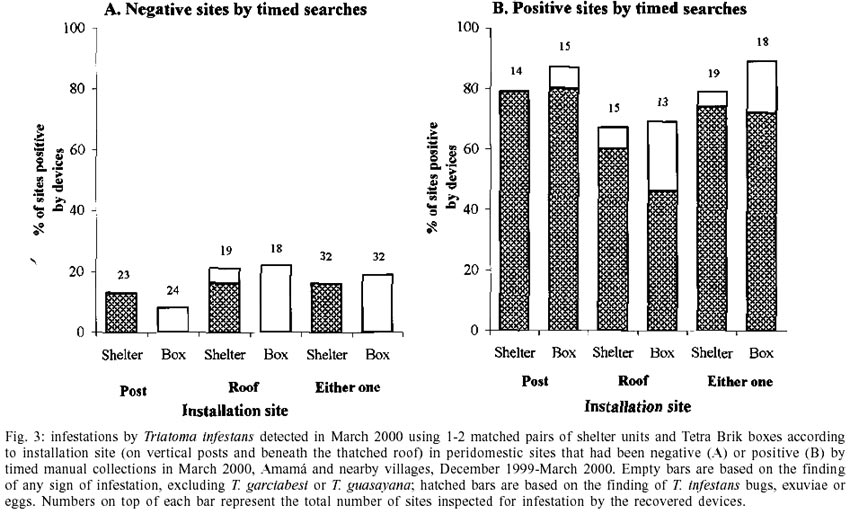
0.50; df = 1; P = 0.48). After a three-month exposure, at least
one of the recovered devices detected some evidence of triatomine infestation
in 89% (boxes) and 79% (shelters) of positive sites, and in 19% (boxes) and
16% (shelters) of negative sites (Fig.
3)(two-tailed binomial tests, P > 0.5).
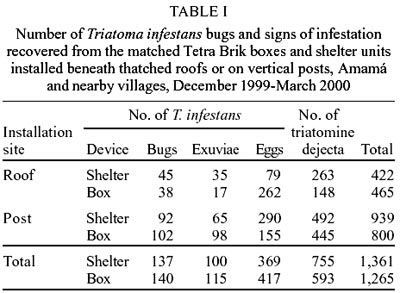
A total of 1,361 and 1,265 signs
of infestation (including 137 and 140 bugs) was recovered from the shelter units
and boxes, respectively (Table I).
The frequency distribution of every sign of infestation in the positive devices
also was significantly different from a random distribution: T. infestans
bugs (c2= 530.9, df =14, P <
0.005), exuviae (c2= 615.6, df
= 28, P < 0.005), eggs (c2=
1676.9, df = 26, P < 0.005) and triatomine dejecta (c2=
3689.35, df = 47, P < 0.005); the variance to mean ratios (14,
18, 24 and 66, respectively) again indicated strong aggregation. The boxes were
as efficient as the shelters in the collection of every sign in either installation
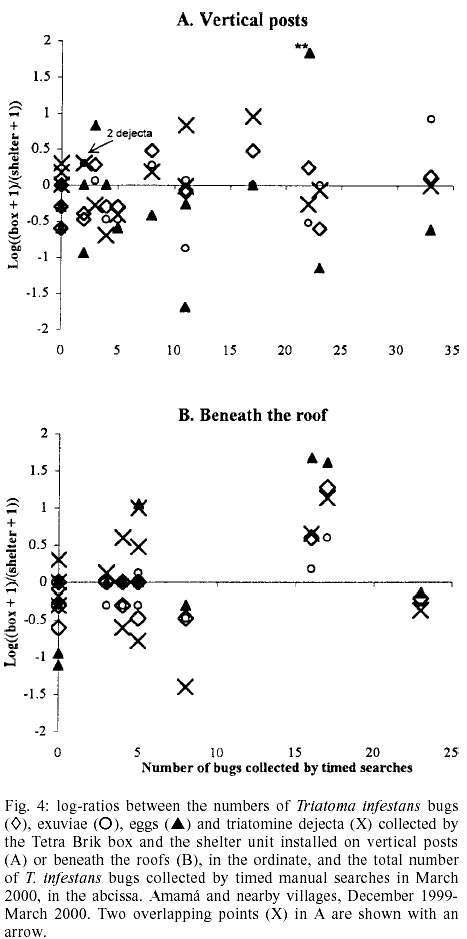
site (Fig. 4). When all signs of
infestation were pooled, the relative sampling efficiency of the devices installed
on the posts (mean log ratio ± 95% C.L., -0.13 ± 0.15) or beneath
the roofs (0.02 ± 0.31) was not significantly different. None of the correlation
coefficients between the log ratios for each type of sign and bug catches by
timed manual collection was statistically significant in either installation
site (for all cases, P > 0.1).
The installation site modified the
sampling efficiency of signs of infestation by shelter units but not by the
boxes (Table I). Shelters installed
on the posts collected significantly more T. infestans bugs (mean log
ratio ± 95% C.L., 0.62 ± 0.39), exuviae (0.40 ± 0.38), eggs (0.57
± 0.42), triatomine dejecta (0.61 ± 0.52), or any sign of infestation
(0.67 ± 0.56) than the paired shelters beneath the roofs. Despite clear
differences at the aggregate level, paired boxes installed on the posts and
beneath the roofs did not differ significantly in the collection of T. infestans
bugs (mean log ratio ± 95% C.L., 0.27 ± 0.98), exuviae (0.22 ±
0.82), eggs (-0.43 ± 1.43), triatomine dejecta (0.37 ± 0.60), or any
sign of infestation (0.25 ± 0.71). The density of triatomine dejecta per
100 cm2 in the languet (median, 5.8; Q1-Q3, 0.8-15) was significantly
larger by at least one order of magnitude than in the accordion (median, 0.4;
Q1-Q3, 0-0.8), wall lining (median, 0.9; Q1-Q3, 0.2-4.7) and the external surface
(median, 0.9; Q1-Q3, 0.4-0.2)(Kruskal-Wallis, c2=
10.73; df = 3; P = 0.013).
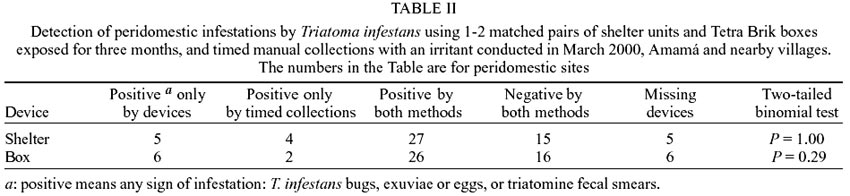
Shelters or boxes exposed for three
months and timed manual collections with an irritant did not differ significantly
in their capacity to detect some evidence of triatomine infestation (Table
II). Both devices and timed searches detected some infestations that were
missed by the alternative method. Results for shelter units and timed collections
in Table II differ from those published
before (Table III in Gürtler et al. 2001b) because the present data series
includes more peridomestic sites exposed for a different and shorter time period
(December to March). The stage structure differed significantly among methods
(c2 = 57.7; df = 2; P
< 0.001). Timed manual collections were biased toward large stages; from
194 bugs collected, 55% were adults, 17% V, 20% III-IV and 8% I-II. In contrast,
the boxes collected 140 bugs, including 14% adults, 19% V, 48% III-IV and 18%
I-II nymphs, whereas the shelter units collected 137 bugs, including 32% adults,
18% V, 22% III-IV, and 29% I-II. In negative sites, T. infestans bugs
were found only in the shelters (6 I, 1 II and 1 V). The devices also detected
29 T. garciabesi bugs and one T. guasayana bug. Spiders, crickets,
small lizards (Gecko sp.), known to prey on triatomines, and cockroaches
were also recovered from the devices.
A previous cost estimate for each
shelter unit was US$1.74 (Gürtler et al. 2001b), or US$0.50 at June 12,
2002 prices. For the box, the current cost estimate is US$0.26, and includes
US$0.14 for materials (leather, corrugated paper and wire) and US$0.12 for 5
min of labor (cutting and assemblage, assuming US$1.43 per labor hour). Additional
expenses related to fieldwork, which for the shelter unit were US$0.40 at 2002
prices, have been reduced to US$0.31 for the box. These include search for a
suitable site and deployment of devices, US$0.12 or 5 min; and inspection and
reinstallation of devices, US$0.19 or 8 min.
DISCUSSION
Tetra Brik boxes were consistently
as sensitive as the shelter units or timed manual collections with an irritant
in detecting foci of T. infestans in a great variety of peridomestic
ecotopes differing in construction materials, resident hosts and intensity of
infestation both in La Rioja and Santiago del Estero Provinces. This is consistent
with previous results (Gürtler et al. 2001b) and supports the replacement
of timed searches or shelter units by boxes for detecting and collecting peridomestic
T. infestans.
Boxes and shelters revealed the presence
of T. infestans bugs in sites where timed manual collections had been
or were negative concurrently. This may be explained by `false negative' results
of timed collections or to recent bug colonization, because the bugs recovered
from the devices installed in negative sites either were first-second instar
nymphs and adults, or late instars and adults. Moreover, in Amamá both
devices detected low-density infestations and other new foci missed by concurrent
timed manual collections, in spite of all searches having been made by the same
highly experienced bug collector. Clearly, rapid timed searches by skilled bug
collectors may fail to detect many peridomestic foci of T. infestans
that, in the absence of treatment, inevitably would increase in abundance and
reinfest the treated areas.
Boxes and shelters apparently provided
similarly suitable conditions that increased the bugs' residence time in the
interior and the likelihood of leaving at least one sign of infestation, which
is consistent with previous results (De Marco et al. 1999, Gürtler et al.
2001b). The aggregation behavior of several triatomine species is mediated by
thigmotaxis and by volatiles present in their feces (Schofield & Paterson
1977, Lorenzo Figueiras et al. 1994). Under laboratory conditions, T. infestans
bugs apparently used feces as chemical marks left just outside their refuges
(Lorenzo & Lazzari 1996). Here we provide the first evidence that this pattern
also occurs in the field, where the density of triatomine feces was significantly
more abundant in the entrances (i.e., the leather languet) than inside the artificial
refuges. Moreover, the large number of triatomine feces found in the devices
could enhance both their attractiveness to T. infestans bugs and the
chance of collecting more bugs, exuviae or eggs. Given the interspecific attraction
of T. infestans bugs to feces of T. garciabesi and T. guasayana
(Lorenzo Figueiras & Lazzari 1998), which also infest peridomestic habitats
in the study area, their feces may further increase the attractiveness of the
artificial refuges for T. infestans.
Inappropriate installation of the
devices may expose them to damage by the resident animals, as occurred in Olta,
or to sunlight and rainfall, or may block the entrances to the bugs. Heavy rainfall
did not negatively affect the materials used in either device or the subsequent
finding of signs of infestation during the dry and wet seasons. It is highly
likely that the Tetra Brik box and leather components may be used for many years,
but the less expensive central cardboard structure may need replacement perhaps
every six months. Tetra Brik boxes that are no longer usable can be recycled
in several ways (available at http://www.tetrapak.com).
The installation site of the shelter
units significantly modified the likelihood of the bugs' leaving some sign of
infestation, perhaps because the units' rigid plastic structure, size and shape
did not fit adequately beneath the roof and may have restricted the access of
bugs. Because the entrances and leather languets of shelters and boxes had the
same size, other factors need to be invoked to explain variations in sampling
efficiency related to installation site. In contrast, the installation site
did not significantly modify the boxes' sampling efficiency of any sign of infestation
in either study area, although the total number of signs collected on the vertical
posts tended to be clearly larger than beneath the roof. We believe that this
was caused by the boxes' effective interception of those bugs that returned
back to the refuges in the thatch.
The box was as sensitive as the shelter
in the collection of every sign of infestation in either installation site at
a wide range of bug abundance. The frequency distribution of each type of sign
of infestation within the boxes and shelters was aggregated, with most devices
bearing few signs and a few devices showing many signs. This overdispersion
may lead to confusion when overall ratios of signs, not ratios of individual
pairs, are used to represent the devices' sampling efficiency. In contrast to
light traps (Schweigmann et al. 1988) and yeast or mammal-baited traps (Lorenzo
et al. 1998, Noireau et al. 1999), both shelters and boxes collected all nymphal
stages and adults of T. infestans, including starved and recently fed
T. infestans bugs (Leonardo A Ceballos et al. unpublished data).
The boxes have several advantages
over the shelter unit regarding simplicity, ease for cutting, adaptability to
the variable surface of wooden poles, and cost. The cost of the shelter units
was reduced by nearly 50% because of the use of recycled milk boxes instead
of plastic, and the reduction of labor time to assemble and set up the boxes
in the field. Compared with the US$17 annual cost of triatomine surveillance
per rural house using timed manual searches with an irritant in Argentina in
the early 1990s (Chuit et al. 1992)(US$4.88 at 12 June 2002 prices), surveillance
of peridomestic sites using boxes is clearly the most cost-effective tool currently
available for large-scale control programs, which are in need to cover some
one million rural houses throughout Argentina. If found acceptable by householders,
the boxes might be incorporated into community-based control programs, in which
the periodic replacement of the central cardboard column could be linked to
the search for evidence of peridomestic infestation by local villagers.
ACKNOWLEDGEMENTS
To Dr Sonia Blanco and her staff
at the National Vector Control Program (Argentina) for providing active support
throughout this study, and to Griseldo Roldán and Isaac Ochoa for expert
assistance during fieldwork. Omar Sitatti kindly provided field accommodation.
Kathe Rogerson kindly provided editorial assistance.
REFERENCES
- Cabrera AL, Willink A 1980. Biogeografía
de América Latina. In Monografía 13, Serie Biología,
Organización de los Estados Americanos, Washington, DC, p. 69-75.
- Canale DM, Cecere MC, Chuit R,
Gürtler RE 2000.Perido-mestic distribution of Triatoma garciabesi
and Triatoma guasayana in north-west Argentina. Med Vet Entomol
14: 383-390.
- Cecere MC, Castañera MB,
Canale DM, Chuit R, Gürtler RE 1999.Trypanosoma cruzi infection
in Triatoma infestans and other triatomines: long term effects of a
control program in rural northwestern Argentina. Pan Am J Publ Health
5: 392-399.
- Cecere MC, Gürtler RE, Canale
D, Chuit R, Cohen JE 1997.The role of the peridomiciliary area in the elimination
of Triatoma infestans from rural Argentine communities. Pan Am J
Publ Health 1: 273-279.
- Chuit R, Paulone I, Wisnivesky-Colli
C, Bo R, Perez AC, Sosa-Stani S, Segura EL 1992. Result of a first step toward
community-based surveillance of transmission of Chagas' disease with appropriate
technology in rural areas. Am J Trop Med Hyg 46: 444-450.
[ Medline
]
- Cohen JE, Gürtler RE 2001.
Modeling household transmission of American trypanosomiasis. Science
293: 694-698. [ Medline
]
- De Marco RJ, Gürtler RE,
Salomón OD, Chuit R 1999. Small-scale field trial of a sensing device
for detecting peridomestic populations of Triatoma infestans (Hemiptera:
Reduviidae) in north-western Argentina. J Med Entomol 36: 884-887.
[ Medline
]
- Dias JCP 1991. Chagas disease
control in Brazil: Which strategy after the attack phase? Ann Soc Belge.
Méd Trop 71 (Suppl. 1): 75-86.
- Dujardin JP, Cardozo L, Schofield
CJ 1996. Genetic analysis of Triatoma infestans following insecticidal
control interventions in central Bolivia. Acta Trop 61: 263-266.
[ Medline
]
- Dujardin JP, Tibayrenc M, Venegas
E, Maldonado L, Desjeux P, Ayala F 1987. Isozyme evidence of lack of speciation
between wild and domestic Triatoma infestans (Heteroptera: Reduviidae)
in Bolivia. J Med Entomol 24: 40-45. [ Medline
]
- Gürtler RE, Cecere MC, Canale
D, Castañera MB, Chuit R, Cohen JE 1999. Monitoring house reinfestation
by vectors of Chagas disease: a comparative trial of detection methods during
a four-year follow-up. Acta Trop 72: 213-234. [ Medline
]
- Gürtler RE, Chuit R, Cecere
MC, Castañera MB 1995. Detecting domestic vectors of Chagas disease:
a comparative trial of six methods in northwest Argentina. Bull WHO
73: 487-494. [ Medline
]
- Gürtler RE, Oneto ML, Cecere
MC, Castañera MB, Canale D 2001a. A simple method to identify triatomine
(Hemiptera: Reduviidae) feces in sensing devices used in vector surveillance
programs. J Med Entomol 38: 147-152. [ Medline
]
- Gürtler RE, Vazquez-Prokopec
GM, Ceballos LA, Lund Petersen C, Salomón OD 2001b.Comparison between
two artificial shelter units and timed manual collections for detecting peridomestic
Triatoma infestans (Hemiptera: Reduviidae) in rural northwestern Argentina.
J Med Entomol 38: 429-436. [ Medline
]
- Lorenzo Figueiras AN, Keningstein
A, Lazzari CR 1994. Aggregation in the haematophagous bug Triatoma infestans:
Chemical signals and temporal pattern. J Ins Physiol 40: 311-316.
- Lorenzo Figueiras AN, Lazzari
CR 1998. Aggregation and interespecific responses in three species of triatominae.
Mem Inst Oswaldo Cruz 93: 133-137.
- Lorenzo MG, Lazzari CR 1996. The
spatial pattern of defaecation in Triatoma infestans and the role of
faeces as a chemical mark of the refuge. J Ins Physiol 42: 903-907.
- Lorenzo MG, Reisenman CE, Lazzari
CR 1998. Triatoma infestans can be captured under natural climatic
conditions using yeast-baited traps. Acta Trop 70: 277-284.
[ Medline
]
- Noireau F, Bastrenta B, Catalá
S, Dujardin JP, Panzera F, Torres M, Perez R, GalvãoC, Jurberg J 2000.
Sylvatic population of Triatoma infestans from the Bolivian Chaco:
from field collection to characterization. Mem Inst Oswaldo Cruz 95 (Suppl.
I):119-122.
- Noireau F, Flores R, Vargas F
1999. Trapping sylvatic triatomi-nae (Reduviidae) in hollow trees. Trans
R Soc Trop Med Hyg 93: 13-14.
- Rabinovich JE, Gürtler RE,
Leal J, Piñero DF 1995. Density estimates of the domestic vector of
Chagas disease, Rhodnius prolixus Ståhl (Hemiptera, Reduviidae),
in rural houses in Venezuela. Bull WHO 73: 347-357.
[ Medline
]
- Schmunis GA, Zicker F, Moncayo
A 1996. Interruption of Chagas' disease transmission through vector elimination.
Lancet 348: 1171.
- Schofield CJ 1978. A comparison
of sampling techniques for domestic populations of Triatominae. Trans R
Soc Trop Med Hyg 72: 449455.
- Schofield CJ, Patterson JW 1977.Assembly
pheromone of Triatoma infestans and Rhodnius prolixus nymphs
(Hemiptera: Reduviidae). J Med Entomol 13: 727-734.
[ Medline
]
- Schweigmann N, Vallvé S,
Muscio O, Guillini M, Alberti A, Wisnivesky-Colli C 1988.Dispersal flight
by Triatoma infestans in an arid area of Argentina. Med Vet Entomol
2: 401-404. [ Medline
]
- Zar JH 1996. Biostatistical
Analysis, 3rd ed., Prentice-Hall, Englewood Cliffs, New Jersey.
- Zeledón R, Rabinovich JE
1981. Chagas disease: an ecological appraisal with special emphasis on its
insect vectors. Ann Rev Entomol 26: 101-133.
Copyright 2002 Instituto Oswaldo
Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02220t1.jpg]
[oc02220f3.jpg]
[oc02220f1.jpg]
[oc02220f2.jpg]
[oc02220t2.jpg]
[oc02220f4.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}