
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 7, 2002, pp. 979-983
|
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(7), October
2002, pp. 979-983
B-cell Infiltration
and Frequency of Cytokine Producing Cells Differ between Localized and Disseminated
Human Cutaneous Leishmaniases
MGS Vieira, F Oliveira/**, S Arruda,
AL Bittencourt*, AA Barbosa Jr, M Barral-Netto/**/***, A Barral /**/***/+
Centro de Pesquisas Gonçalo
Moniz-Fiocruz, Rua Valdemar Falcão 121, 40295-001 Salvador, BA, Brasil
*Serviço de Anatomia Patológica, Hospital Professor Edgard Santos
**Faculdade de Medicina, Universidade Federal da Bahia, Salvador, BA, Brasil
***Instituto de Investigação em Imunologia, Instituto do Milênio,
São Paulo, SP, Brasil
+Corresponding author. Fax: +55-71-356.4320. E-mail:
abarral@cpqgm.fiocruz.br
FO received a scientific initiation fellowship and ALB, MB-N and AB are
CNPq senior investigators.
This work was supported
by Pronex (CNPq, Brazil) and Papes -Fiocruz.
Received 8 February 2002
Accepted 29 July 2002
Code Number: oc02221
Biopsies from human localized
cutaneous lesions (LCL n = 7) or disseminated lesions (DL n = 8) cases were
characterized according to cellular infiltration,frequency of cytokine (IFN-g,
TNF-a) or iNOS enzyme producing cells. LCL,
the most usual form of the disease with usually one or two lesions, exhibits
extensive tissue damage. DL is a rare form with widespread lesions throughout
the body; exhibiting poor parasite containment but less tissue damage. We demonstrated
that LCL lesions exhibit higher frequency of B lymphocytes and a higher intensity
of IFN-g expression. In both forms of the
disease CD8+ were found in higher frequency than CD4+
T cells. Frequency of TNF-a and iNOS producing
cells, as well as the frequency of CD68+ macrophages, did not differ
between LCL and DL. Our findings reinforce the link between an efficient control
of parasite and tissue damage, implicating higher frequency of IFN-g
producing cells, as well as its possible counteraction by infiltrated B cells
and hence possible humoral immune response in situ.
Key words: leishmaniasis - B lymphocytes
- cytokines - CD8+ T cells - IFN-g
Cutaneous leishmaniasis is a worldwide
disease with severe deformating potential in new world. It affects preferentially
young economically active patients representing a large burden to the public
health system in developing countries. Protection against all forms of leishmaniasis
is dependent on cell-mediated immunity (CMI), but the contribution of some cells
and cytokines in human disease deserves further scrutiny.
CD8+T cells have been
implicated in protection (Muller et al. 1991) being high IFN-g
producers in a murine model of leishmaniasis (Chan 1993). Their role seems to
be more in the secondary than in the primary immune response (Muller et al.
1993, 1994). On the other hand, the course of leishmaniasis in mice lacking
beta 2-microglobulin (beta 2-m) gene did not differ from their wild-type counterparts
(Overath & Harbecke 1993, Wang et al. 1993, Huber et al. 1998) lessening
a role of antigen presentation by major histocompatibility complex class I (MHC
I) molecules. In man, a higher percentage of CD8+ over CD4+
T cells was found in mucocutaneous leishmaniasis (MCL) lesions (Castes
& Tapia 1998), compared to localized cutaneous lesions (LCL), although similar
distributions of CD4+ and CD8+ in LCL have been reported
(Barral et al. 1987, Esterre et al. 1992, Lima et al. 1994). The presence of
cytotoxic CD8+ T cells has been reported in peripheral blood of MCL
but not in LCL patients (Brodskyn et al. 1997). Expansion of CD8+
T cells occurs in the peripheral blood of individuals vaccinated against leishmaniasis
(Mendonça et al. 1995, Gurunathan et al. 2000). Especially, the percentage
of activated CD8+ T cells was higher in fast responding than in slow
responding volunteers to vaccination (Pompeu et al. 2001).
The role of B cells in leishmaniasis
is also not clear. High antibody levels are present in the more severe clinical
form of the cutaneous disease, namely diffuse cutaneous leishmaniasis (DCL)
(Schurr et al. 1986, Mengistu et al. 1990), but B cell depletion does not alter
the susceptibility or resistance pattern to Leishmania infection in mice
(Babai et al. 1999, Brown & Reiner 1999). It seems that B cells are important
to induce anti-Leishmania CD4+ Th1 cells and DTH reaction,
in the resistant mouse strain, and take part in the humoral response development
in susceptible animals (Scott & Farrell 1982, Scott et al. 1986).
Predominance of Th1 cytokines like
IFN-g, IL-12, IL-2 and TNF-a
over Th2 cytokines, IL-4, IL-5 IL-10 and TGF-b, is
correlated in mice to the resistance profile against Leishmania infection
(Belosevic et al. 1989, Chatelain et al. 1992, Lezama-Davila et al. 1992, Barral
et al. 1993). Imunological studies in humans demonstrated a combination of Th1
and Th2 cytokines with predominance of Th1 in MCL, Th2 predominate in DCL and
predominance of Th1 profile in LCL patients (Caceres-Dittmar et al. 1993, Castes
et al. 1993, Tapia et al. 1993).
Human tegumentary leishmaniasis has
a diversity of clinical presentations. Evaluating the in situ immune response
in different presentations of human leishmaniasis may help in defining the role
of cells and cytokines in the course of disease. Herein we report on a comparison
of LCL to DL. LCL is the typical presentation, where the majority of the patients
have one or two ulcerated lesions, elevated borders and necrotic center, preferentially
localized at the lower limbs. LCL has a clinical course of several months, but
may exhibit spontaneous healing and clinical cure. Disseminated leishmaniasis
(DL) is a rare condition (1% of cutaneous leishmaniasis patients) and is characterized
by the presence of multiple (> 20 lesions) ulcerated lesions in several parts
of the body. Despite the larger number of lesions, DL patients respond promptly
to antimonial treatment and heal faster than LCL patients. The mechanism of
dissemination is not clear but the rapid onset, lack of lymph node enlargement
and the presence of fever and chills suggest hematogenic dissemination (Costa
et al. 1986, Carvalho et al. 1994). Both LCL and DL patients exhibit anti-Leishmania
CMI (Carvalho et al. 1994). Differences in the in situimmune reactions of
these two forms of human leishmaniasis may help us elucidating the participating
mechanisms in the effective response against the parasite.
MATERIALS AND METHODS
Biopsy - Biopsies were obtained
from seven patients with LCL and eight cases of DL, all from the endemic area
Corte de Pedra, Bahia, Brazil with predominance of L. braziliensis (Barretto
et al. 1981). LCL patients presented unique lesions with a necrotic center and
elevated borders. All biopsies were taken from the borders of ulcers. DL patients
showed multiple lesions, varying from papules, acneiform lesions and few ulcers.
Only acneiform lesions were biopsy from DL patients. Biopsies in these, cases,
involved the whole lesion. Characteristics of the patient population are summarized
in the Table. Diagnosis of leishmaniasis
were based on clinical and pathological observations and confirmed both parasitologically
(presence of Leishmania amastigotes in tissue sections) and immunologically
(a delayed type hypersensitivity – test larger than 5 mm and/or anti-Leishmania
IgG antibody titers above 1/16).
Immunohistochemical reactions
- Formaline-fixed and paraffin-embedded 4 µm sections were incubated
in hot humid vapor of pH 6.0 citrate buffer for 5 min. An additional incubation
with 10% non-fat milk for 20 min at room temperature was used to block unspecific
reactions. The primary antibodies and dilutions used were: CD3 (1:100; DAKO
Corporation, Carpinteria, CA, USA), CD4 (1:10; DAKO), CD8 (1:20; DAKO), CD20
(1:100; KAKO) and CD68 (1:20; DAKO). Following primary antibody incubation (40
min, at 37°C), the sections were reacted with biotinilated anti-rabbit
antibody (1:300; DAKO) or pork anti-rabbit antibody (1:600; DAKO), followed
by peroxidase-conjugated streptavidin (1:50; DAKO) for 30 min at 37°C.
Diaminebenzidine (Vector Inc., Burlingame, CA, USA) was used as chromogen. For
cytokine staining, endogenous peroxidase was blocked by incubation with 3% H2O2
for 20 min at room temperature, followed by an incubation with 3% of trypsin
(Sigma-Aldrich, St. Louis, MO, USA) and an additional incubation with 0.1% saponin
(Sigma-Aldrich) for 15 min for permeabilizatiom. Blocking of unspecific reactions
was performed with 2% normal goat serum for 20 min. The primary antibodies used
were anti-IFN-g (1:200; Genzyme Corporate Offices,
Cambridge, England), anti-TNF-a (1:200; Genzyme Corporate
Offices) and anti-iNOS (1:500, Genzyme Corporate Offices). Tonsil sections were
used as positive controls for cell characterization and sections from pulmonary
tuberculosis granulomatous lesions as controls for cytokines and iNOS staining.
Monoclonal antibodies were substituted for non-immune rabbit immunoglobulins
or irrelevant mouse antibodies as negative controls.
Evaluation criteria - The
numbers of positive cells for each marker was counted in five 400X fields comprising
a total dermic sectional area of 1.4 mm2, using an image analysis
and processing system (Quantimet Q500MC; Leica, Cambridge, UK). Reactivity to
cytokines or iNOS was graded based on the number of positive cells out of 100
cells observed, and classified as negative – no cells positive; low –
up to 20% of positive cells; moderate – 21% to 50% of positive cells; or
high if more than 50% of the cells were positive.
Statistical analysis - Comparisons
of cell numbers (CD4+and CD8+cells) in each disease presentation
were made by Wilcoxon paired non-parametric test. For the comparison between
leishmaniasis presentations (LCL x DL for each parameter), the unpaired nonparametric
Mann-Whitney test was used. Significance was determined as p < 0.05. All
statistical tests and graphs are done with Prism-GraphPad version 3 (GraphPad
Software Inc., San Diego, CA, USA).
RESULTS
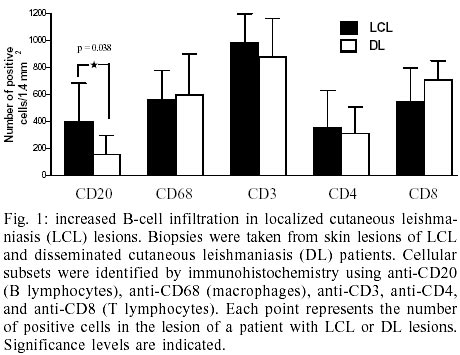
Cellular infiltration - T
cells were the most frequent cell in both LCL and DL lesion infiltrate comprising
approximately 34.8% of the total number of cells. A large number of CD68+
macrophages were also observed (21.6%) followed by CD20+ B cells
on the range of 10.3% (Fig. 1). The
numbers of total T cells and macrophages did not differ between LCL and DL lesions
but there was a statistically significant higher frequency of B lymphocytes
in LDL lesions (Fig. 1).
T cells subsets - In order
to explore the predominant T cell subset present in the lesion we evaluated
CD4 and CD8 expression at LCL and DL lesions. There was a predominance of CD8+
T cells in both disease presentations. CD8+ T cells represented 68.9%
of the T cells in LCL whereas CD4+ T cells comprised 31.1% (Fig.
1). Similarly in DL there were 69.4% CD8+ cells versus 30.6%
CD4+ T cells (Fig. 1).
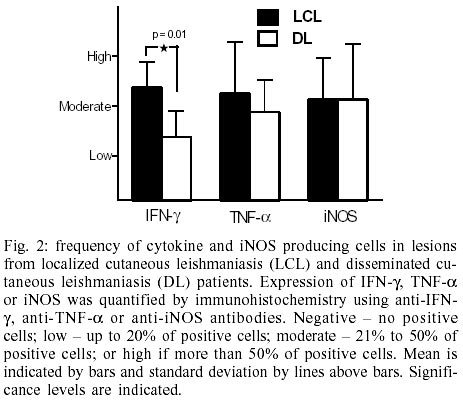
Frequency of cell producing cytokines
and iNOS enzyme at the lesion site - Expression of IFN-g
was significantly more intense in LCL samples than in DL (Fig.
2). In the 8 samples examined in LCL, IFN-gexpressing
cells represented 30% of the total cells, whereas in DL less than 20% of the
total cells expressing IFN-g. No differences were
observed between LCL and DL regarding iNOS and TNF-a
expression.
DISCUSSION
Lesions from LCL patients exhibited
a higher frequency of IFN-g producing cells than
DL, which may suggest more efficient protection since it is the main cytokine
implicated in Leishmania killing and host protection in all forms of
leishmaniasis (Sadick et al. 1986, Belosevic et al. 1989, Sadick et al. 1990).
The higher expression of IFN-g in LCL may result
in parasite containment at the site of the sand fly bite, preventing parasite
dissemination. Such higher expression is not due to a larger cellular infiltration
since the numbers of T lymphocytes as well as their CD4+ and CD8+
subsets, were similar between LCL and DL lesions. It is possible that the frequency
of activated T cells differs between these two conditions, and other unexplored
elements such as IL-10 or TGF-b expression may be
contributing to a lesser frequency of IFN-g producing
cells in DL patients.
Despite differences in IFN-g,
there was no difference in the frequency of iNOS enzyme expressing cells between
LCL and DL lesions. It is possible that other factors also able of inducing
iNOS expression, like TNF-a (Liew et al. 1990a,b),
compensate for the diminished IFN-g in DL, resulting
in a similar expression of iNOS. NO is considered a key element to Leishmania
killing in the murine models of leishmaniasis (Green, Crawford et al. 1990,
Green, Meltzer et al. 1990, Liew et al. 1990), but the NO involvement in human
protection against leishmaniasis has never been clearly demonstrated. Our finding
of a similar frequency of iNOS enzyme producing cells in two largely diverse
presentations of human leishmaniasis with different levels of parasite control
gives support to the idea that NO is not elemental in Leishmania killing
in man.
CD8+ T cells were more
frequent than CD4+ in both LCL and DL lesions at our observations.
The role these cells play in human leishmaniasis is unclear. They might participate
directly in the immune response against the parasite, and secrete Th1 cytokines,
principally IFN-g. However, our results do not lend
support to a prominent role of CD8+ T cells in human leishmaniasis,
as two diverse forms displayed similar levels of these cells. Future studies
may elucidate the role of these cells by double staining of activated cells
and intracellular cytokine in situ.
Our results show B cell infiltration
higher in LCL than in DL lesions. Besides antibody production, these cells have
been implicated in driving Th response, antigen presentation, and expression
of costimulator molecules (Liu et al. 1995, van Essen et al. 1995, Amigorena
& Bonnerot 1998, Brown & Reiner 1999). In LCL lesions a higher IFN-g
expression could be stimulating B cells to secrete antibodies like IgG1, that
could be involved in opsonization and more efficient phagocitosis, leading to
better parasite containment. However parasites may use this way to gain entrance
into phagocytes, helping in perpetuating the lesion. At present we have no sufficient
elements to clarify the complex role of B cells in human cutaneous leishmaniasis.
Immune response in leishmaniasis
is implicated in both protection and immunopathology. The predominant IFN-g
and B cell infiltration at LCL, which may be operative in competent containment
of the parasites, may also contribute to tissue injury, which might lead to
the larger and persistent lesions observed in LCL as compared to DL patients.
Additionally, tissue damage may also impair access of the drugs to lesion site,
which might be related to a less efficient response to drug treatment observed
in LCL patients.
ACKNOWLEDGMENTS
To the expert assistance provided
by the technicians of the Serviço de Anatomia Patológica from
Hospital Universitário Professor Edgard Santos, Universidade Federal
da Bahia and the CPqGM-Fiocruz.
REFERENCES
- Amigorena S, Bonnerot C 1998.
Role of B-cell and Fc receptors in the selection of T-cell epitopes. Curr
Opin Immunol 10: 88-92. [ Medline
]
- Babai B, Louzir H, Cazenave PA,
Dellagi K 1999. Depletion of peritoneal CD5+ B cells has no effect on the
course of Leishmania major infection in susceptible and resistant mice.
Clin Exp Immunol 117: 123-129. [ Medline
]
- Barral A, Barral-Netto M, Yong
EC, Brownell CE, Twardzik DR, Reed SG 1993. Transforming growth factor beta
as a virulence mechanism for Leishmania braziliensis. Proc Natl
Acad Sci USA 90: 3442-3446. [ Medline
]
- Barral A, Jesus AR, Almeida RP,
Carvalho EM, Barral-Netto M, Costa JM, Badaro R, Rocha H, Johnson WD 1987.
Evaluation of T-cell subsets in the lesion infiltrates of human cutaneous
and mucocutaneous leishmaniasis. Parasite Immunol 9: 487-497.
[ Medline
]
- Barretto AC, Cuba CA, Marsden
PD, Vexanat JA, De Belder M 1981. Epidemiological features of American cutaneous
leishmaniasis in an endemic region of the state of Bahia, Brazil. I. Human
leishmaniasis. Bol Oficina Sanit Panam 90: 415-424.
- Belosevic M, Finbloom DS, Van
Der Meide PH, Slayter MV, Nacy CA 1989. Administration of monoclonal anti-IFN-gamma
antibodies in vivo abrogates natural resistance of C3H/HeN mice to infection
with Leishmania major. J Immunol 143: 266-274.
- Brodskyn CI, Barral A, Boaventura
V, Carvalho E, Barral-Netto M 1997. Parasite-driven in vitro human lymphocyte
cytotoxicity against autologous infected macrophages from mucosal leishmaniasis.
J Immunol 159: 4467-4473. [ Medline
]
- Brown DR, Reiner SL 1999. Polarized
helper-T-cell responses against Leishmania major in the absence of
B cells. Infect Immun 67: 266-270. [ Medline
]
- Caceres-Dittmar G, Tapia FJ, Sanchez
MA, Yamamura M, Uyemura K, Modlin RL, Bloom BR, Convit J 1993. Determination
of the cytokine profile in American cutaneous leishmaniasis using the polymerase
chain reaction. Clin Exp Immunol 91: 500-505. [ Medline
]
- Carvalho EM, Barral A, Costa JM,
Bittencourt A, Marsden P 1994. Clinical and immunopathological aspects of
disseminated cutaneous leishmaniasis. Acta Trop 56: 315-325.
[ Medline
]
- Castes M, Tapia FJ 1998. Immunopathology
of American tegumentary leishmaniasis. Acta Cient Venez 49:
42-56. [ Medline
]
- Castes M, Trujillo D, Rojas ME,
Fernandez CT, Araya L, Cabrera M, Blackwell J, Convit J 1993. Serum levels
of tumor necrosis factor in patients with American cutaneous leishmaniasis.
Biol Res 26: 233-238. [ Medline
] [ Lilacs
]
- Costa JM, Marsden PD, Llanos-Cuentas
EA, Netto EM, Carvalho EM, Barral A, Rosa AC, Cuba CC, Magalhaes AV, Barreto
AC 1986. Disseminated cutaneous leishmaniasis in a field clinic in Bahia,
Brazil: a report of eight cases. Am J Trop Med Hyg 89: 319-323.
- Chan MM 1993. T cell response
in murine Leishmania mexicana amazonensis infection: production of
interferon-gamma by CD8+ cells. Eur J Immunol 23: 1181-1184.
[ Medline
]
- Chatelain R, Varkila K, Coffman
RL 1992. IL-4 induces a Th2 response in Leishmania major-infected mice.
J Immunol 148: 1182-1187. [ Medline
]
- Esterre P, Dedet JP, Frenay C,
Chevallier M, Grimaud JA 1992. Cell populations in the lesion of human cutaneous
leishmaniasis: a light microscopical, immunohistochemical and ultrastructural
study. Virchows Arch A Pathol Anat Histopathol 421: 239-247.
[ Medline
]
- Green SJ, Crawford RM, Hockmeyer
JT, Meltzer MS, Nacy CA 1990. Leishmania major amastigotes initiate
the L-arginine-dependent killing mechanism in IFN-gamma-stimulated
macrophages by induction of tumor necrosis factor-alpha. J Immunol
145: 4290-4297. [ Medline
]
- Green SJ, Meltzer MS, Hibbs Jr
JB, Nacy CA 1990. Activated macrophages destroy intracellular Leishmania
major amastigotes by an L-arginine-dependent killing mechanism. J Immunol
144: 278-283. [ Medline
]
- Gurunathan S, Stobie L, Prussin
C, Sacks DL, Glaichenhaus N, Iwasaki A, Fowell DJ, Locksley RM, Chang JT,
Wu CY, Seder RA 2000. Requirements for the maintenance of Th1 immunity in
vivo following DNA vaccination: a potential immunoregulatory role for CD8+
T cells. J Immunol 165: 915-924. [ Medline
]
- Huber M, Timms E, Mak TW, Rollinghoff
M, Lohoff M 1998. Effective and long-lasting immunity against the parasite
Leishmania major in CD8-deficient mice. Infect Immun 66:
3968-3970. [ Medline
]
- Lezama-Davila CM, Williams DM,
Gallagher G, Alexander J 1992. Cytokine control of Leishmania infection
in the BALB/c mouse: enhancement and inhibition of parasite growth by local
administration of IL-2 or IL-4 is species and time dependent. Parasite
Immunol 14: 37-48. [ Medline
]
- Liew FY, Li Y, Millott S 1990a.
Tumor necrosis factor-alpha synergizes with IFN-gamma
in mediating killing of Leishmania major through the induction of nitric
oxide. J Immunol 145: 4306-4310. [ Medline
]
- Liew FY, Li Y, Millott S 1990b.
Tumour necrosis factor (TNF-alpha) in leishmaniasis.
II. TNF-alpha-induced macrophage leishmanicidal
activity is mediated by nitric oxide from L-arginine. Immunology 71:
556-559.
- Liew FY, Millott S, Parkinson
C, Palmer RM, Moncada S 1990c. Macrophage killing of Leishmania parasite
in vivo is mediated by nitric oxide from L-arginine. J Immunol 144:
4794-4797. [ Medline
]
- Lima HC, Vasconcelos AW, David
JR, Lerner EA 1994. American cutaneous leishmaniasis: in situ characterization
of the cellular immune response with time. Am J Trop Med Hyg 50:
743-747. [ Medline
]
- Liu Y, Wu Y, Ramarathinam L, Guo
Y, Huszar D, Trounstine M, Zhao M 1995. Gene-targeted B-deficient mice reveal
a critical role for B cells in the CD4 T cell response. Int Immunol
7: 1353-1362. [ Medline
]
- Mendonça SC, De Luca PM,
Mayrink W, Restom TG, Conceição-Silva F, Da-Cruz AM, Bertho
AL, Da Costa CA, Genaro O, Toledo VP 1995. Characterization of human T lymphocyte-mediated
immune responses induced by a vaccine against American tegumentary leishmaniasis.
Am J Trop Med Hyg 53: 195-201. [ Medline
]
- Mengistu G, Akuffo HO, Yemane-Berhan
T, Britton S, Fehniger TE 1990. Serum antibody specificities to Leishmania
aethiopica antigens in patients with localized and diffuse cutaneous leishmaniasis.
Parasite Immunol 12: 495-507. [ Medline
]
- Muller I, Kropf P, Etges RJ, Louis
JA 1993. Gamma interferon response in secondary Leishmania major infection:
role of CD8+ T cells. Infect Immun 61: 3730-3738. [
Medline
]
- Muller I, Kropf P, Louis JA, Milon
G 1994. Expansion of gamma interferon-producing CD8+ T cells following secondary
infection of mice immune to Leishmania major. Infect Immun 62:
2575-2581. [ Medline
]
- Muller I, Pedrazzini T, Kropf
P, Louis J, Milon G 1991. Establishment of resistance to Leishmania major
infection in susceptible BALB/c mice requires parasite-specific CD8+ T cells.
Int Immunol 3: 587-597. [ Medline
]
- Overath P, Harbecke D 1993. Course
of Leishmania infection in beta 2-microglobulin-deficient mice. Immunol
Lett 37: 13-17. [ Medline
]
- Pompeu MM, Brodskyn C, Teixeira
MJ, Clarencio J, Van Weyenberg J, Coelho IC, Cardoso SA, Barral A, Barral-Netto
M 2001. Differences in gamma interferon production in vitro predict the pace
of the in vivo response to Leishmania amazonensis in healthy volunteers.
Infect Immun 69: 7453-7460. [ Medline
]
- Sadick MD, Heinzel FP, Holaday
BJ, Pu RT, Dawkins RS, Locksley RM 1990. Cure of murine leishmaniasis with
anti-interleukin 4 monoclonal antibody. Evidence for a T cell-dependent, interferon
gamma-independent mechanism. J Exp Med 171: 115-127.
[ Medline
]
- Sadick MD, Locksley RM, Tubbs
C, Raff HV 1986. Murine cutaneous leishmaniasis: resistance correlates with
the capacity to generate interferon-gamma in response to Leishmania antigens
in vitro. J Immunol 136: 655-661. [ Medline
]
- Schurr E, Kidane K, Yemaneberhan
T, Wunderlich F 1986. Cutaneous leishmaniasis in Ethiopia: I. Lymphocyte transformation
and antibody titre. Am J Trop Med Parasitol 37: 403-408.
- Scott PA, Farrell JP 1982. Experimental
cutaneous leishmaniasis: disseminated leishmaniasis in genetically susceptible
and resistant mice. Am J Trop Med Hyg 31: 230-238. [
Medline
]
- Scott P, Natovitz P, Sher A 1986.
B lymphocytes are required for the generation of T cells that mediate healing
of cutaneous leishmaniasis. J Immunol 137: 1017-1021.
[ Medline
]
- Tapia FJ, Caceres-Dittmar G, Sanchez
MA, Fernandez AE, Convit J 1993. The cutaneous lesion in American leishmaniasis:
leukocyte subsets, cellular interaction and cytokine production. Biol Res
26: 239-247. [ Medline
] [ Lilacs
]
- van Essen D, Kikutani H, Gray
D 1995. CD40 ligand-transduced co-stimulation of T cells in the development
of helper function. Nature 378: 620-623. [ Medline
]
- Wang ZE, Reiner SL, Hatam F, Heinzel
FP, Bouvier J, Turck CW, Locksley RM 1993. Targeted activation of CD8 cells
and infection of beta 2-microglobulin-deficient mice fail to confirm a primary
protective role for CD8 cells in experimental leishmaniasis. J Immunol
151: 2077-2086. [ Medline
]
Copyright 2002 Instituto Oswaldo
Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02221f1.jpg]
[oc02221t1.jpg]
[oc02221f2.jpg]
|

{kind=link}
{kind=link}
{kind=link}