
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 7, 2002, pp. 991-995
|
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(7), October
2002, pp. 991-995
Complete Nucleotide
Sequence Analysis of a Brazilian Dengue Virus Type 2 Strain
Flávia Barreto dos Santos/+,
Marize Pereira Miagostovich, Rita Maria Ribeiro Nogueira, Dianna Edgil*,
Hermann Gonçalves Schatzmayr, Lee W Riley*, Eva Harris*
Departamento de Virologia, Instituto
Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
*Division of Infectious Diseases, School of Public Health, University of California,
Berkeley, CA, USA
+Corresponding author. Fax: + 55-21-2598.4373.
E-mail: Flaviab2001@aol.com
This work was supported
by grants from the Fogarty International Center of the National Institutes of
Health (TW-00905), Conselho Nacional de Desenvolvimento Científico e
Tecnológico (CNPq- 40.0164/98-1) and Fundação Oswaldo Cruz.
Received 13 March 2002
Accepted 9 May 2002
Code Number: oc02223
In the last decade, dengue fever
(DF) in Brazil has been recognized as an important public health problem, and
an increasing number of dengue haemorrhagic fever (DHF) cases have been reported
since the introduction of dengue virus type 2 (DEN-2) into the country in 1990.
In order to analyze the complete genome sequence of a DEN-2 Brazilian strain
(BR64022/98), we designed primers to amplify contiguous segments of approximately
500 base pairs across the entire sequence of the viral genome. Twenty fragments
amplified by reverse transcriptase-PCR were cloned, and the complete nucleotide
and the deduced amino acid sequences were determined. This constitutes the first
complete genetic characterization of a DEN-2 strain from Brazil. All amino acid
changes differentiating strains related to the Asian/American-Asian genotype
were observed in BR64022/98, indicating the Asiatic origin of the strain.
Key words: dengue virus type 2 -
complete genome - Brazil
Dengue (DEN) viruses are responsible
for the most important arthropod-borne viral diseases in humans in terms of
morbidity and mortality. Due to global population growth, increased urbanization,
and the spread of both the mosquito vector and the four DEN serotypes, dengue
is a major emerging problem in tropical and subtropical areas worldwide. Nowadays
it is estimated that more than 100 million cases of dengue fever (DF) and 250,000
cases of the more severe form of the disease, dengue hemorrhagic fever/dengue
shock syndrome (DHF/DSS), occur annually (Halstead 1997).
The four serotypes of DEN (DEN-1
to -4), genus Flavivirus (family Flaviviridae), consist of a single-stranded
positive-sense RNA genome of approximately 10,700 bases in length, surrounded
by a nucleocapsid and a lipid envelope containing the envelope and membrane
proteins. The genome contains a single open reading frame that encodes three
structural proteins designated C (core protein), M (membrane protein), and E
(envelope protein) and seven nonstructural proteins (NS1, NS2A, NS2B, NS3, NS4A,
NS4B and NS5), which is flanked by a 5' and a 3' nontranslated (NTR) region
(Chambers et al. 1990, Lindenbach & Rice 2001).
The partial sequencing of various
regions of the DEN genome has been employed to determine the genetic variation
of dengue viruses and to characterize genotypes within each serotype (Rico-Hesse
1990, Blok et al. 1991, Deubel et al. 1993, Lewis et al. 1993, Lanciotti et
al. 1994, 1997, Chungue et al. 1995, Shurtleff et al. 2001). Genotypic characterization
has been useful in molecular epi-demiological studies that monitor the distribution
of the circulating genotypes in endemic areas (Rico- Hesse et al. 1997, 1998,
Messer et al. 2001).
Phylogenetic analysis of DEN-2 strains
initially identified five genotypes, including two of Southeast Asian origin
(Asian and American-Asian) (Rico-Hesse 1990, Lewis et al. 1993); further studies
incorporating additional strains resulted in the merging of these two subgroups
into one Asian/American-Asian genotype (Rico-Hesse et al. 1998). Detailed analysis
of a 240-base pair (bp) sequence spanning the E/NS1 junction in a collection
of Latin American strains showed that the Asian/American-Asian DEN-2 variant
had been introduced into the Americas and had spread through the region over
the past two decades, replacing the native American genotype (Rico-Hesse et
al. 1997). This Asian/American-Asian genotype has been associated with DHF cases,
whereas the native American genotype has to date only been associated with classic
DF. Recently, Leitmeyer et al. (1999) sequenced the full-length genome of DEN-2
viruses directly from viremic sera and identified nucleotide differences that
consistently differentiated the Asian/American-Asian DEN-2 viruses from the
native American DEN-2 strains.
Since the introduction of DEN-2 into
Brazil resulted in an increase in disease severity and in the number of epidemics
(Nogueira et al. 1993, Zagne et al. 1994, Souza et al. 1995, Vasconcelos et
al. 1995), we cloned and determined the complete nucleotide and the deduced
amino acid sequences of a Brazilian DEN-2 strain isolated during an epidemic
that occurred in 1998. This constitutes the first complete genetic characterization
of a DEN-2 Brazilian strain since its introduction into the country in 1990
(Nogueira et al. 1990).
MATERIALS AND METHODS
Viral strain - The DEN-2 strain
analyzed in this study was obtained from the collection of the Flavivirus Laboratory,
Department of Virology, Instituto Oswaldo Cruz-Fiocruz. The strain was isolated
in 1998 from the serum of a DF patient by inoculation into Aedes albopictus
C6/36 cell line (Igarashi 1985), and the serotype was identified by immunofluorescence
using type-specific monoclonal antibodies (Gubler et al. 1984). The second passage
was used for amplification, cloning and sequencing.
Primer design - Synthetic
oligonucleotide primer pairs were designed to amplify overlapping fragments
of approximately 500 bp spanning the complete DEN-2 genome based on an alignment
of the full-length sequences of representative Asian/American-Asian genotype
DEN-2 strains. The sequences of the oli-gonucleotide primers used for the reverse
transcriptase-PCR (RT-PCR) are available upon request. The sequences of the
strains Jamaica 1409/83 (accession no. M20558), New Guinea C/44 (AF038403),
Thailand 16681/64 (U87411), and Thailand K10010/94 (AF100460) were retrieved
from the National Center for Biomedical Investigation (NCBI) database, and the
sequence alignments were generated using GeneJockey Software (Biosoft, Inc.,
Ferguson, MO).
RNA extraction and RT-PCR amplification
- Viral RNA was extracted from the supernatant of infected cells, and fragments
were amplified using a one-tube procedure according to Harris et al.
(1998). The RT-PCR mixture contained 50 mM KCl, 10 mM Tris (pH 8.5), 0.1% Triton
X-100, 0.01% gelatin, each of the four deoxynucleotide triphosphates at a concentration
of 200 µM, 1.5 mM MgCl2, 5 mM dithiotreitol, each primer at
a final concentration of 10 µM, 0.025 U of RAV-2 (Amershan Corporation,
Arlington Heights, IL) and 0.025 U of Taq DNA Polymerase (AmpliTaq,
Perkin-Elmer Corporation, Foster City, CA) per ml of reaction. Five microliters
of the extracted RNA were reverse transcribed at 42°C for 60 min, followed
directly by 40 cycles of amplification consisting of 94°C for 30 sec, 60°-65°C
for 1 min, and 72°C for 2 min, with a final extension at 72°C for
10 min. Amplification was conducted using a Model 2400 thermal cycler (Perkin-Elmer,
Norwalk, CT) or PTC-200-60 thermocycler (MJ Research, Inc., Watertown, MA).
To sequence the DEN-2 virus genome, cDNA fragments amplified by PCR were cloned
into TA vector pCR 2.1 (Invitrogen, San Diego, CA) according to the manufacturer's
protocol. The sequence between genome positions 1 to 99 and 9974 to 10637 was
determined by direct sequencing of the PCR product.
Sequencing of recombinant polypeptides
clones - Plasmids containing the recombinant DEN genes were sequenced in
both directions using the BigDye Terminator Cycle Sequencing Ready Reaction
kit (Applied Biosystems, Foster City, CA), 3.2 pmol of T7 forward (AATACGACTCACTATAG)
and M13 Reverse (AACAGCTATGACCATG) primers combined with 0.5 µg of plasmid
DNA. Thermocycling conditions consisting of 25 cycles of 96oC for
30 s, 50oC for 1 min and 60oC for 4 min were used as recommended
by the manufacturer. After purification using 3 M sodium acetate and 70% ethanol,
the DNA was dried in a vacuum centrifuge. The pellet was resuspended in 20 µl
of template suppression reagent, heated for 2 min at 95oC and kept
on ice until 10 µl was loaded on an Applied Biosystems Prism 310 sequencer
using perfomance-optimized polymer 6 (Applied Biosystems).
Sequence and phylogenetic analysis
- The sequence analysis of the clones was performed by using GeneJockey
Software (Biosoft, Inc.). Nucleotide and amino acid identity was determined
using BLAST (http://www.ncbi.nlm.nih.gov/Entrez/).
Secondary structure predictions were performed using MFOLD (http://bioinfo.math.rpi.edu/~mfold/rna/form1.cgi).
Phylogenetic trees were constructed using PAUP* software (Sinauer Associates,
Inc., Sunderland, MA) using the maximum parsimony algorithm, with a heuristic
search and tree bisection-reconnection branch-swapping. Representative sequences
from DEN serotypes 1, 3, and 4 were used as an outgroup to root the trees. The
bootstrap method, with 100 replications, was used to estimate the reliability
of the predicted trees.
RESULTS
To obtain the full-length sequence
of a low-passage Brazilian DEN-2 strain, we designed primers to amplify contiguous
segments across the entire sequence of the BR64022/98 virus genome. Twenty fragments
obtained by RT-PCR were cloned into the TA vector and sequenced in both directions.The
sequences between genome positions 1 and 99 (5'NTR) and 9,974 to 10,637 (3'NTR)
were determined by directly sequencing the RT-PCR product in both directions.
The complete sequence of BR64022/98 strain is 10,637 nucleotides long and was
submitted to NCBI under accession number AF489932.
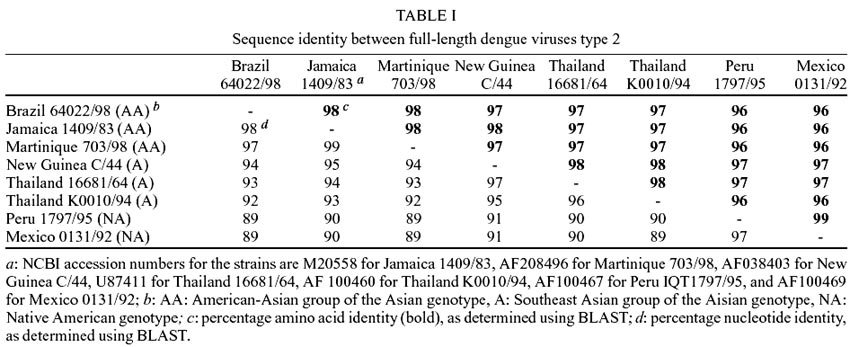
The overall homology at the nucleotide
(nt) and amino acid (aa) level between the Brazilian strain and representatives
of the Asian/American-Asian genotype and the native American genotype is shown
in Table I. The Brazilian strain
was most closely related to the American-Asian strains (98% nt, 98% aa identity),
followed by the Southeast Asian strains (92% nt, 97% aa), and finally the native
American genotype (89% nt, 96% aa).
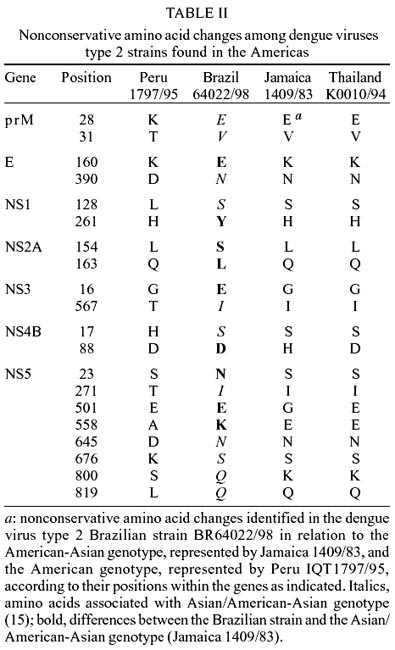
All amino acid changes reported to
differentiate strains related to the Southeast Asian genotype from the native
American genotype were observed in BR64022/98 (Table
II), indicating the Asiatic origin of our strain. There were a number of
additional nonconserved amino acid substitutions in the Brazilian strain relative
to the Asian/American-Asian strain, notably charge reversals in E (K160E), NS4B
(H88D), and NS5 (E558K) and charge additions in NS3 (G16E) and NS5 (G501E).
The sequences of the 5' and 3' NTRs of the Brazilian strain, and thus the secondary
structure predictions, are identical to those of the American-Asian viruses
(e.g., Jamaican 1409/83 strain).
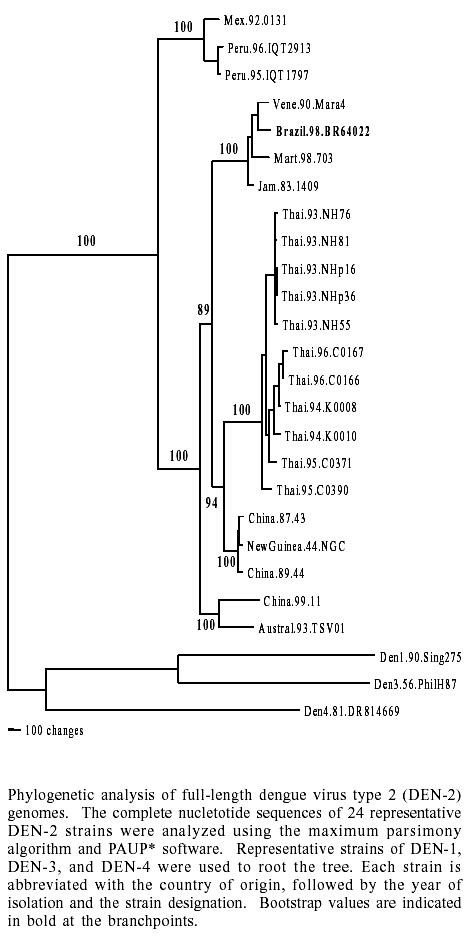
Figure
shows the phylogenetic analysis performed using the maximum parsimony algorithm
of BR64022/98 and full-length DEN-2 sequences retrieved from the NCBI database.
The tree placed the Brazilian strain in the American-Asian group of the DEN-2
Asian genotype, represented here by Jamaica 1409/83, Martinique 703/98, and
Venezuela Mara4/90 strains. Similar phylogenetic trees were generated using
maximum likelihood and neighbor joining algorithms (data not shown).
DISCUSSION
Full-length analysis of the nucleotide
and amino acids sequences of the Brazilian strain showed that it was more closely
related to Asian/American-Asian DEN-2 genotype than to the native American DEN-2,
confirming the Asiatic origin of the Brazilian strain. These data corroborate
the results previously obtained by the partial sequencing of different genomic
fragments (Deubel et al. 1993, Rico-Hesse et al. 1997, Miagostovich et al. 1998).
A direct association between the introduction of Asian DEN-2 virus with severe
disease in the Americas has been shown previously (Rico-Hesse et al. 1997),
and our results are consistent with this scenario. The direction of transmission
from Southeast Asia to the Americas has been demonstrated as well, since DEN-2
viruses from Brazil, Colombia, Mexico and Venezuela have a common progenitor
with those from Southeast Asian (Rico-Hesse et al. 1997).
Comparing the complete genomic sequences
of DEN-2 strains from the native American and Asian genotypes, Leitmeyer et
al. (1999) identified candidate nucleotide differences responsible for the virulent
genotype. All the amino acids characteristic of the Asian genotype were identified
in the Brazilian strain BR64022/98. These similarities with the Asian genotype
were also reported recently for the Martinique 703/98 strain after analysis
of the complete genome (Tolou et al. 2000). The Asian-specific non-conserved
amino acid differences, as well as the additional differences specific to the
Brazilian strain, are found in E, NS3, and NS5. Changes in E could affect immunogenicity
or cell entry/tropism, whereas changes in NS3 (helicase/protease) and NS5 (RNA-dependent
RNA polymerase) could affect replication efficiency. In addition, differences
in the predicted secondary structure of the 5' and 3' NTRs are found between
the two genotypes; in these regions, the Brazilian isolate was identical to
the American-Asian strains in sequence and consequently in the predicted secondary
structures. The NTRs presumably mediate interaction with cellular and viral
factors that regulate viral translation and RNA replication (Lindebach &
Rice 2001).
In Brazil, DEN-2 was introduced initially
in the state of Rio de Janeiro in 1990, and after a short period of time, its
presence was detected in 24 of 26 states (Nogueira et al. 1990, Ministry of
Health, Brazil, http://www.funasa.gov.br).
From 1986 to 2001, about 2,000,000 cases of DF were reported. DHF/DSS cases
(928 to date) were only reported after the introduction of DEN-2 in 1990, when
both DEN-1 and DEN-2 co-circulated. The appearance of the first DHF/DSS cases
and the increase in the number of hospitalizations resulting from DEN-2 secondary
infection corroborate data that suggest an increase in virulence of this strain
(Nogueira et al. 1990, 1993, Zagne et al. 1994, Rico-Hesse et al. 1997). Although
secondary infection with a distinct serotype increases the risk of experiencing
the severe form of the disease (Monath 1994, Halstead 1997), the intrinsic properties
of dengue virus strains themselves are also important in determining the severity
of the disease (Rosen 1977). Studies performed in Peru, where only the native
American DEN-2 genotype circulates, showed that even with high secondary infection
rates, no DHF/DSS cases were observed (Watts et al. 1999).
The partial sequencing of the junction
from E/NS1 and analysis by restriction site specific-PCR of DEN-2 Brazilian
strains from the latest epidemics (2000-2001) have shown that this Asian/American-Asian
genotype is still the only one circulating in Brazil (data not published). Molecular
epidemiological studies continue with strains isolated from both DF and DHF/DSS
cases in order to detect the eventual introduction of a new DEN-2 genotype and
evaluate its effect of the epidemiology of the disease.
ACKNOWLEDGEMENTS
To Sangwei Lu for technical advice,
Marcia Firmani and Josefina Coloma for assistance with sequencing, Karen Clyde
for secondary structure predictions, and to Rebeca Rico-Hesse and Brian Brunelle
for help with phylogenetic analysis.
REFERENCES
- Blok J, Gibbs AJ, McWilliam SM,
Vitaran UT 1991. NS1 gene sequences from eight dengue-2 viruses and their
evolutionary relationships with other dengue-2 viruses. Arch Virol
118: 209-233. [ Medline
]
- Chambers TJ, Hahn CS, Galler R,
Rice CM 1990. Flavivirus genome organization, expression, and replication.
Annu Rev Microbiol 44: 649-688. [ Medline
]
- Chungue E, Cassar O, Drouet MT,
Guzman MG, Laille M, Rosen L, Deubel V 1995. Molecular epidemiology of dengue-1
and dengue-4 viruses. J Gen Virol 76: 1877-1884.
- Deubel V, Nogueria RMR, Drouet
MT, Zeller H, Reynes JM, Ha DQ 1993. Direct sequencing of genomic cDNA fragments
amplified by the polymerase chain reaction for molecular epidemiology of dengue-2
viruses. Arch Virol 129: 197-210. [ Medline
]
- Gubler DJ, Kuno G, Sather E, Valez
M, Oliver A 1984. Mosquito cell cultures and specific monoclonal antibodies
in surveillance for dengue viruses. Am J Trop Med Hyg 33: 158-165.
[ Medline
]
- Halstead SB 1997. Epidemiology
of dengue and dengue hemorrhagic fever. In DJ Gubler, G Kuno (eds), Dengue
and Dengue Hemorrhagic Fever, CAB International, New York, p. 23-44.
- Harris E, Roberts TG, Smith L,
Selle J, Kramer LD, Valle S, Sandoval E, Balmaseda A 1998. Typing of dengue
viruses in clinical specimens and mosquitoes by single-tube multiplex reverse
transcriptase-PCR. J Clin Microbiol 36: 2634-2639. [ Medline
]
- Igarashi A 1985. Mosquito cell
cultures and the study of arthropod-borne togaviruses. Adv Virus Res 3:
21-39.
- Lanciotti RS, Gubler DJ, Trent
DW 1997. Molecular evolution and phylogeny of dengue-4 viruses. J Gen Virol
78: 2279-2286.
- Lanciotti RS, Lewis JG, Gubler
DJ, Trent DW 1994. Molecular evolution and epidemiology of dengue-3 viruses.
J Gen Virol 75: 65-75.
- Leitmeyer KC, Vaughn DW, Watts
DM, Salas R, Villalobos de Chacon I, Ramos C, Rico-Hesse R 1999. Dengue virus
structural differences that correlate with pathogenesis. J Virol
73: 4738-4747. [ Medline
]
- Lewis JG, Chang G-J, Lanciott,
RS, Kinney RM, Mayer LM, Trent DW 1993. Phylogenetic relationships of dengue-2
viruses. Virology 197: 216-224. [ Medline
]
- Lindenbach BD, Rice CM 2001. Flaviviridae:
the viruses and their replication. In DM Knipe, PM Howley (eds), Fields
Virology, Lippincott Williams and Wilkins, Philadelphia, p. 991-1041.
- Messer WB, Harris E, Gubler DJ,
Sivananthan K, Preethimala LD, de Silva AM 2001. The molecular evolution and
epidemiology of dengue 3, subtype III. Am J Trop Med Hyg 65: 282.
- Miagostovich MP, Nogueira RMR,
Schatzmayr HG, Lanciotti RS 1998. Molecular epidemiology of DEN-2 virus in
Brazil. Mem Inst Oswaldo Cruz 93: 625-626. [ Medline
] [ SciELO
]
- Monath TP 1994. Dengue: the risk
to developed and developing countries. Proc Natl Acad Sci USA 91:
2395-2400. [ Medline
]
- Nogueira RMR, Miagostovich MP,
Lampe E, Schatzmayr HG 1990. Isolation of dengue virus type 2 in Rio de Janeiro.
Mem Inst Oswaldo Cruz 85: 253.
- Nogueira RMR, Miagostovich MP,
Lampe E, Souza RW, Zagne SMO, Schatzmayr HG 1993. Dengue epidemic in the state
of Rio de Janeiro, Brazil, 1990-1: co-circulation of dengue 1 and dengue 2
serotypes. Epidemiol Infect 111: 163-170.
- Rico-Hesse R 1990. Molecular evolution
and distribution of dengue viruses type 1 and 2 in nature. Virology 174:
479-493. [ Medline
]
- Rico-Hesse R, Harrison LM, Nisalak
A, Vaughn DW, Kalayanarooj S, Green S, Rothman AL, Ennis FA 1998. Molecular
evolution of dengue type 2 virus in Thailand. Am J Trop Med Hyg
58: 96-101. [ Medline
]
- Rico-Hesse R, Harrison LM, Salas
RA, Tovar D, Nisalak A, Ramos C, Boshell J, de Mesa MT, Nogueira RM, da Rosa
AT 1997. Origins of dengue type 2 viruses associated with increased pathogenicity
in the Americas. Virology 230: 244-251. [ Medline
]
- Rosen L 1977. The Emperor's New
Clothes revisited, or reflections on the pathogenesis of dengue hemorrhagic
fever. Am J Trop Med Hyg 26: 337-43. [ Medline
]
- Shurtleff AC, Beasley DWC, Chen
JJY, Ni H, Suderman MT, Wang H, Xu R, Wang E, Weaver SC, Watts DM, Russell
KL, Barrett ADT 2001. Genetic variation in the 3' non-coding region of dengue
viruses. Virology 281: 75-87. [ Medline
]
- Souza RV, Cunha RV, Miagostovich
MP, Timbó MJ, Montenegro F, Pessoa ETFP, Nogueira RMR, Schatzmayr HG
1995. An outbreak of dengue virus infection in the State of Ceará,
Brazil. Mem Inst Oswaldo Cruz 90: 345-346.
- Tolou H, Couissinier-Paris P,
Mercier V, Pisano MR, De Lamballerie X, De Micco P, Durand JP 2000. Complete
nucleotide sequence of dengue type 2 virus from the French West Indies. Biochem
Biophys Res Commun 277: 89-92.
- Vasconcelos PFC, Menezes DB, Melo
LP, Paula Pessoa ETF, Rodrigues SG, Travassos Da Rosa ES, Timbó MJ,
Coelho ICB, Montenegro F, Travassos da Rosa, JFS, Andrade FMO, Travassos da
Rosa APA 1995. A large epidemic of dengue fever with dengue hemorrhagic cases
in Ceará State, Brazil, 1994. Rev Inst Med Trop São Paulo
37: 252-255.
- Watts DM, Porter KR, Putvatana
P, Vazquez B, Calampa C, Hayes CG, Halstead SB 1999. Failure of secondary
infection with American genotype dengue 2 to cause dengue haemorrhagic fever.
Lancet 354: 1431-1434.
- Zagne SMO, Alves VGF, Nogueira
RMR, Miagostovich MP, Lampe E, Tavares W 1994. Dengue haemorrhagic fever in
the state of Rio de Janeiro, Brazil: a study of 56 confirmed cases. Trans
R Soc Trop Med Hyg 88: 677-679. [ Medline
]
Copyright 2002 Instituto Oswaldo
Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02223t2.jpg]
[oc02223f1.jpg]
[oc02223t1.jpg]
|

{kind=link}
{kind=link}
{kind=link}