
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 97, Num. 7, 2002, pp. 1049-1055
|
Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(7), October
2002, pp. 1049-1055
Dynamics of Evolution
and Resistance to Starvation of Triatoma vitticeps (Stal 1859) (Reduviidae:
Triatominae), Submitted to Two Different Regimens of Food Deprivation
Carlos José de Carvalho Moreira+,
Maria Celeste Dias Spata
Núcleo de Interação
Trypanosoma cruzi-Vetores da Doença de Chagas, Laboratório
de Doenças Parasitárias, Departamanto de Medicina Tropical, Instituto
Oswaldo Cruz-Fiocuz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
+Corresponding author. Fax:
+55-21-2280.3740. E.mail: moreira@ioc.fiocruz.br
Received 28 November 2001
Accepted 29 July 2002
Code Number: oc02232
The resistance to starvation of
Triatoma vitticeps has been analyzed comparatively, according to different
regimens of food deprivation under laboratory conditions. One cohort, composed
of 100 specimens of each evolutionary nymphal stage, was submitted to continuous
fasting until death; the second group, arranged in the same way, was fed once
on chicken. Through this work, it was possible not only to compare the results
obtained from the first group with other authors' results but, also, to analyze
the dynamics of evolution, molting, longevity and the increase in insect longevity,
in the second group. The average values recorded for survival time revealed
statistical differences between the two groups. Among the important results
detected, there is one that deserves to be emphasized: the incredible increase
in longevity among insects that received only one feed – an average increase
in survival time that reached 2.95 to 3.30 times in nymphs of 3rd and 4th stages,
respectively. One 5th stage nymph survived for up to 350 days and the females
may triplicate their survival rate, what represents an important epidemiological
factor. The knowledge about this type of biological characteristic of T.
vitticeps may contribute to prevent the domiciliation of this species, what
seems to be incipient in some municipal districts, in Brazil.
Key words: Triatoma vitticeps
- life cycle - resistance in starvation - triatomine longevity - Brazil
Triatominae are insects belonging
to the Hemiptera Order, Family Reduviidae, and Subfamily Triatominae. Nowadays,
there are about 131 vector species known, distributed among 6 tribus and 17
genera (Carcavallo et al. 2001). All of them are absolute bloodsuckers, regardless
of age, sex or evolutionary stage. They are susceptible to infections by Trypanosoma
cruzi (Chagas, 1909) and acquire the infection after feeding from an infected
mammal, thus becoming vectors of Chagas disease, which today affects thousands
of people, mainly in South America.
Triatoma vitticeps (Stal,
1859) is found in four Brazilian states: Bahia, Minas Gerais, Espírito
Santo and Rio de Janeiro. Basically, it is still considered a wild species for
two reasons: (a) the high number of adult insects captured in human dwellings
infected with the T. cruzi (Santos et al. 1969b, Silveira et al. 1983,
Ferreira et al. 1986, Sessa & Carias 1986, Dias et al. 1989); (b) the low
rates of fortuitous infections in humans, in these areas (Santos et al. 1969
a, c, Pinto et al. 1969, 1986, Barros et al. 1975, Peçanha et al. 1983,
Silveira et al. 1983, Dias et al. 1989). This indicates that the infection is
acquired outside the residences, in places that have already been described
as natural biotopes. However, Santos et al. (1969b) had already found domestic
specimens in the State of Espírito Santo, where, at the occasion, there
was a suspicion that the disease transmission had occurred through the action
of the insect vector.
Silveira et al. (1984) registered
the occurrence of this species in 21 cities of the State of Espírito
Santo. In two towns, Cachoeiro do Itapemirim and Guarapari, the process of domiciliation
was confirmed by the finding of household nymphs.
In the State of Rio de Janeiro, Ferreira
et al. (1986), with the collaboration of the Superintendence of Public Health
Campaigns, identified the presence of T. vitticeps in 4 municipal districts
of the northern area (Campos, Itaocara, Porciúncula and São João
da Barra); 3 of the metropolitan area (Maricá, Niterói and Rio
de Janeiro) and 2 of the highland area (Santa Maria Madalena and São
Sebastião do Alto). Eighteen adult insects among the 51 tested presented
cruzi-like flagellated forms. In the municipal district of Santa
Maria Madalena, 8 household nymphs were collected.
Recently, Gonçalves et al.
(1998) captured 465 specimens of T. vitticeps (98% were captured indoors),
in an eco-epidemiological study performed in the municipal district of Santa
Maria Madalena. From this total, 202 (111 alive and 91 dead) were examined and
infection by Trypanosoma cruzi wasconfirmed in 88 live bugs and in 31
of the dead ones.
This species had already demonstrated
to be suitable as an experimental model for xenodiagnostic tests (Perlowagora-Szumlewicz
et al. 1990); this means that it behaves as a proper vector species, presenting
high infectivity rates and good parasitic density, for periods of up to 120
days after the infection, when infected by 7 different strains of T. cruzi.
It was also tested in endemic areas of Chagas disease, in the State of Minas
Gerais (Moreira & Perlowagora-Szumlewicz 1997), with better results than
T. infestans (Klug, 1834). During that study, it was also verified a
high level of resistance demonstrated by the low mortality rate. However, other
characteristics are important to define a good vectorial species, as for the
defecation timing during or after feeding, which affects the possibility of
transmission of T. cruzi to man (Gonçalves et al. 1988).
The domiciliation process of T.
vitticeps seems to be due to environmental modifications of those areas,
as the increase of residences and concomitant shortage of insects' alimentary
sources, condition already detected by Barreto (1976) and Silveira et al. (1983).
However, there are still few pertinent data concerning the biology of this species
and the present work intends to contribute with more information, through original
observations, that may help in the efforts to avoid a possible domiciliation
of this species.
MATERIALS AND METHODS
All the insects used in this study
were derived from our laboratory colony, maintained in the Núcleo de
Interação T. cruzi-Vetores da Doença de Chagas,
Departamento de Medicina Tropical, Instituto Oswaldo Cruz, Fundação
Oswaldo Cruz.
Cohorts of 100 specimens of each
nymphal stage recently emerged were submitted to continuous fast until death.
More than 100 specimens of each evolutionary
stage were fed only one time on chicken (Gallus gallus), and 100 well
engorged specimens were separated, forming another cohort. This second group
was separated according to the feeding date, so that it should be observed the
evolution dynamic of the nymphs as well as the mean capacity of blood ingestion
per stage.
The bugs were kept in 235 mm x 655
mm glass cylinders, covered with cheesecloth. The bottom of the cylinders were
covered with a circle of filter paper (K40) and, in their interior, there was
an accordion-pleated strip of the same paper, in order to increase the area
of contact of the insects and to remove the excess of humidity, in all cases
of the insects were fed a single time. The adult insects and those ones in the
5th nymph instar were divided between two glass jars, to avoid overcrowding.
All groups were daily checked for
mortality and ecdysis.
During the accomplishment of the
experiment, the bugs were maintained at controlled environmental temperature
and humidity, with averages of 26°C ± 2°C and 75% ± 5%,
respectively.
For statistical comparisons, we used
t Student test.
RESULTS
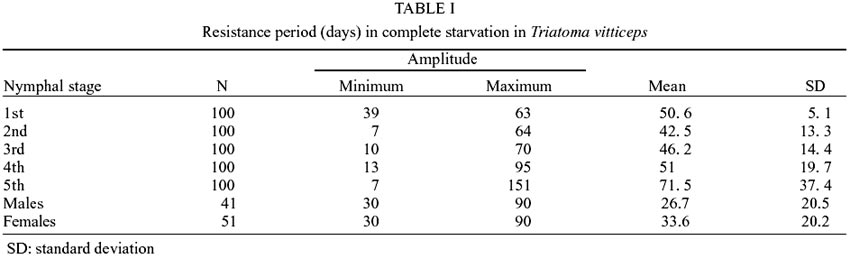
The increasing of survival time is
proportional to the nymphal stage maintained under conditions of complete fasting
until death. Nymphs of the first stage presented the largest pre-mortality period:
39 days. Adult insects presented a lower resistance average, as compared to
other evolutionary stages: 26.7 and 33.6 days for males and females, respectively
(Table I). In Table
I it is also demonstrated that the averages varied in an increasing manner,
except for the 1st stage, which presented a superior average (50.6) as compared
to the two following stages. The shortest resistance was observed in the 2nd
and 5th stages (7 days) and the longest, in the 5th stage (151 days).
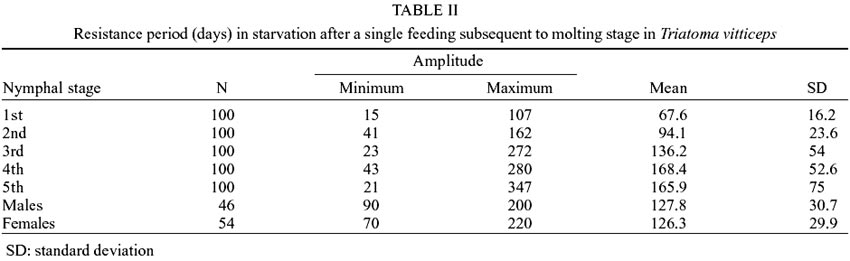
Table
II shows results obtained when nymphs were maintained in starvation, after
a single feeding after molting, for each evolutionary stage. The survival average
rate increased, according to the stage of development until the 4th stage, at
which point the rate was found to be a marginally superior than at the 5th stage
(168.4 and 165.9, respectively). The shortest resistance was observed at the
1st stage (15 days), in contrast to fasting nymphs, and the longest one was
once again verified in nymphs at the 5th stage (347 days). All differences among
the averages presented in Tables I
and II, tested separately for each
stage, using the t-test of Student (p < 0.05), are statistically significant.
The percentage of molted nymphs (Table
III) varied from 39% to 75% at the 4th and 2nd stages, respectively.
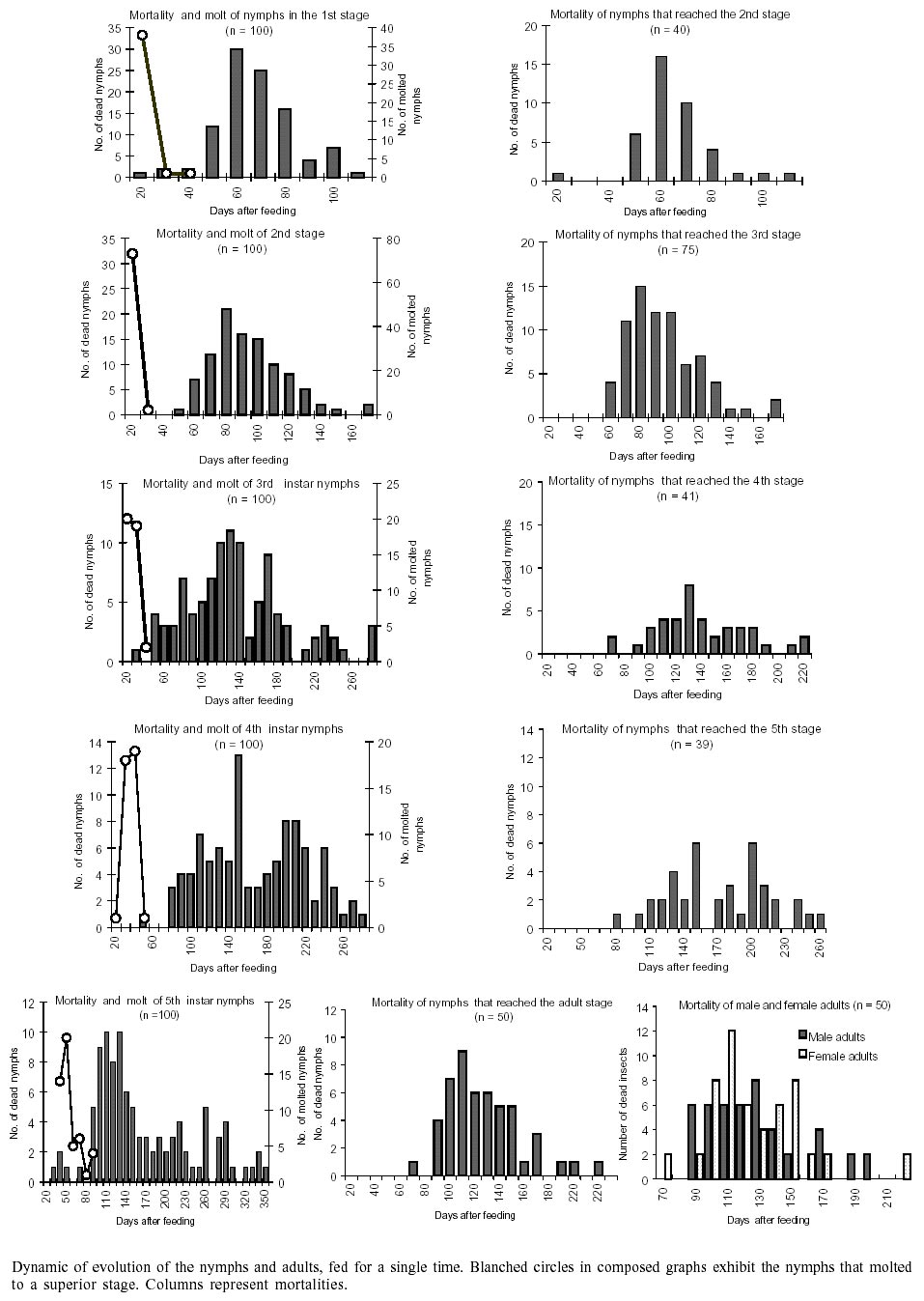
The dynamics of evolution, mortality
and molting of nymphs and adults fed for a single time also varied in agreement
with nymphal stages (Figure). The
time of survival of nymphs that molted to a subsequent stage, as compared to
those that stayed at the initial stage of development (in the selection of insects
for this study), can be also verified in the Figure.
This period was slightly lower in nymphs at 4th stage (proceeding from the 3rd
stage) and at the adult stage (proceeding from the 5th stage), compared with
nymphs that did not move on to subsequent stages. However, at the 5th stage
this difference was more accentuated (350 vs 220 days).
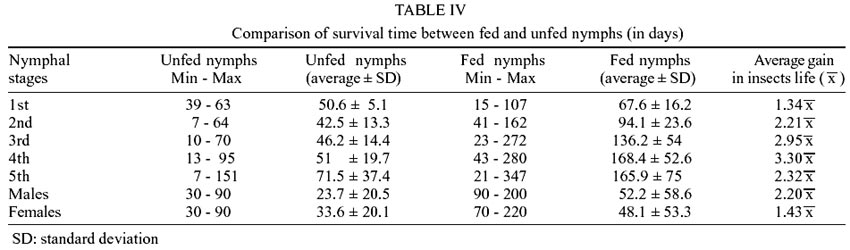
Table
IV shows, in a comparative way, the longevity differences between nymphs
that were fed a single time and those maintained under conditions of fasting
until death. The major longevity difference among the last live specimen registered
for each group was observed at the 3rd (202 days) and 5th stages (196 days).
At the 5th stage, this difference is of greater importance, since this stage
precedes the adult phase.
In this study, the average gain in
the lifetime of the insects turned the differences statistically significant
(p < 0.05), in comparison with the nymphs maintained in total starvation.
The increase in longevity was proportional to the increase in the developmental
stages.
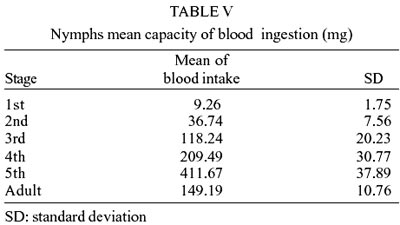
The nymphs mean capacity for blood
ingestion is shown in Table V. Likewise
other already analyzed parameters, the capacity of blood ingestion showed a
continued increase until the 5th stage.
DISCUSSION
According to Neiva and Lent (1936),
the first reports concerning the capacity to resist conditions of fasting observed
in insects were made by Charles Darwin, who noticed a specimen that resisted
lack of food for four months. Leboubène observed a still longer period
and Porter registered a specimen of T. infestans that survived for 17
months (apud Galvão et al. 1996).
Uribe (1926) reported the survival
of one 3rd stage nymph of Rhodnius prolixus Stal, 1859 during 5 months
and, since then, several authors have described the fasting capacity of these
insects. Pellegrino (1952) emphasized the epidemiological importance of this
characteristic in Triatominae and, according to this author, the resistance
to starvation provides the passive transport of the insect to another place
and its subsequent development, so that it soon finds a favorable environment
for such. It also represents an escape mechanism against the action of insecticides,
which are applied in endemic areas.
According to Dias (1965) and Perlowagora-Szumlewicz
(1969), this resistance might be of great importance and directly affect the
control campaigns against these vectors, since domesticated species can hide
in places, like wall openings, that are inaccessible to the action of insecticides
or to its possible residual effects (the insects would be protected from the
action of insecticides during the time that the residual effect of the product
lasted); thus, the possibility of re-colonization is increased.
In this work, the resistance averages
found were lower than those reported by Silva (1985) and Gonçalves et
al. (1988). The exceptions were the averages found by those authors in nymphs
of the 1st stage, maintained under starvation. These different results were
probably influenced by laboratory conditions, such as different blood sources
supplied and variations in environmental conditions (humidity and temperature).
The highest averages were reported by Gonçalves et al. (1989), who used
mice as alimentary source.
In our study, adult insects demonstrated
a lower resistance in comparison with other stages. In this last stage the female
insect was more resistant than the male (33.6 and 23.7 days, respectively).
Data reported by Silva (1985) and Gonçalves et al. (1989) demonstrate
alternating results: sometimes higher for the male and sometimes higher for
the female.
The averages observed in the 2nd
and 3rd instar nymphs were also inferior to the value observed in the 1st stage
(Table I). This result was influenced
either by food reserves carried over from the eggs, that are maintained by nymphs
throughout their entire life, until death, as by the premature mortality of
some nymphs (6 nymphs died during the first 10 days, at the second stage, and
2 nymphs died on the 10th and 20th days, at the 3rd stage). Precocious mortality
as observed in nymphs at the 2nd stage was also observed by Galvão et
al. (1996), when they studied the resistance to starvation in T. nitida Usinger,
1939; by Cortez and Gonçalves (1998) for T. rubrofasciata De Geer,
1773; and by Martinez-Ibarra and Katthain-Duchateau (1999) for T.
pallidipenis Stal, 1872. In our case, it seems to be due mainly to the incapacity
to ingest the blood meal, since dead bugs had no alimentary contents in their
digestive tracts.
Another important aspect was the
evolution process of nymphs fed only once in each stage (Figure).
This kind of study begins to be valued by some authors, and not only the study
of resistance to absolute fast, because this factor associated with the capacity
of the insects to feed from several alimentary sources constitute an important
survival strategy. In this context, Dias-Lima and Sherlock (2001) performed
a comparative study on resistance to starvation, after a single blood meal by
stage, in six species of vectors. They verified different resistance rates for
different evolutionary stages, drawing attention to T. matogrossensis (Leite
and Barbosa, 1953), which resisted up to 575 days, at the 4th nymph stage.
According to Galvão et al.
(1995) some triatomine species can reach the 4th stage with just
one feeding in each stage, which is in agreement with the findings of Brasileiro
and Perondini (1974) results with T. brasiliensis Neiva, 1911. On the
other hand, Heitzmann-Fontenelle (1972) and Gonçalves et al. (1997),
both working with T. pseudomaculata Corrêa & Espínola,
1964, demonstrated the requirement for more than one feeding at each stage for
the successful development of this species. The present results indicate that
the percentage of molted nymphs varied from 39% to 75%, at 4th and 2nd stages,
respectively (Table III).
The largest rate of nymphs that molted
to a superior evolutionary stage was observed in nymphs at the second stage,
followed by 5th stage nymphs.
The time of survival in the cohorts
of selected nymphs was longer than of those that passed through ecdysis, for
nymphs at the same stages (the first ones belonging to the previously separated
cohorts at the beginning of experiments and the last ones originating from ecdysis),
which can be attributed to the metabolic costs associated with the process of
ecdysis. This can be observed in Figure
comparing, for example, the graph that shows mortality in nymphs that reached
the 2nd stage (n = 40) with the graph that demonstrates the mortality and molting
of nymphs at the same stage (n = 100), and so forth. These differences can also
be explained by the larger quantities of blood ingested by the nymphs that stayed
at the initial stage, at the moment that the cohorts were created (Table
V).
An important result was the average
increase in survival rates for insects that fed only a single time (Table
IV). These insects presented an average gain of survival time that varied
from 1.34 times (1st stage) to 4.79 times (males). The females may triplicate
their survival rate. Similar date were seen with nymphs at the 3rd and 4th stages,
what represents another important epidemiological factor. The great survival
capacity demonstrated by the 5th instar nymphs where the last insect died within
350 days, after a single feeding, must be emphasized. Thirty nine percent of
the nymphs fed at the 4th stage passed on to the 5th stage (Figure)
and that, in this last stage, 10% of the nymphs survived 275 days after the
single feeding. If their survivors manage to feed at this stage, 50% of them
will pass on to the adult phase and will survive long enough to reproduce (10%
of the adults can survive up to 160 days).
Lima et al. (1991) tested experimentally
the toxicity of several synthetic insecticides (hexachlorocyclohexane, DDT,
triclorfon, malathion, cypermetrin, deltametrin and permetrin). She added them
to the food intake of six triatomine species (Panstrongylus megistus, T.
infestans, T. pseudomaculata, T. vitticeps, T. brasiliensis and
Rhodnius prolixus) and, among all these species, T. vitticeps demonstrated
the largest level of resistance to any tested insecticide, except for HCH in
concentration of 1 g/l, in spite of the fact that T. vitticeps is not
a household species. This could indicate that this species is naturally resistant.
According to Silva et al. (2001),
T. vitticeps is invading residences also in the State of Minas Gerais, where
it is sometimes lodged, also presenting high infection rates by T. cruzi.
In spite of the fact that a human being seldom is infected by the parasite,
its circulation is happening in reservoirs and there is risk for becoming endemic,
by linking the wild cycle to the domestic one.
The resistance to insecticides and
resistance to fasting after a single blood meal at each nymph stage, as the
present work has demonstrated, may increase the risk of domiciliation of T.
vitticeps in some areas. Therefore, we conclude that an epidemiological
policy is vital in those areas under risk, mainly because of the inevitable
environmental changes caused by man. Hence if the necessary actions to control
these insects are needed, the knowledge of the time that the species may resist
to starvation after a single blood meal be of importance for elaboration of
such actions, thus avoiding flaws in the methodology.
ACKNOWLEDGEMENTS
To Dr Teresa Cristina M Gonçalves,
for her critical review of the manuscript.
REFERENCES
- Barreto MP 1976. Possible role
of wild mammals and triatomines in the transmission of Trypanosoma cruzi
to man. In International Symposium on New Approaches in American Trypanosomiasis
Research, Belo Horizonte, MG, 1975. Proceedings Washington DC, Pan American
Health Organization, p. 307-316 (PAHO-Scientific Publication 318).
- Barros GC, Mayrink W, Salgado
AA, Barros RCG, Sessa PA 1975. Contribuição para o conhecimento
da doença de Chagas autóctone no Espírito Santo. Rev
Inst Med Trop São Paulo 17: 319-329. [ Medline
]
- Brasileiro VLF, Perondini ALP
1974. Biologia de T. brasiliensis (Hemiptera, Reduviidae, Triatominae)
I. Tempo de sucção e repleção de ninfas de 4°
e 5° estádios. Rev Bras Ent 18: 43-50.
- Carcavallo RU, Jurberg J, Lent
H, Galvão C, Steindel M, Pinto CJC 2001. Nova espécie do complexo
oliveirai (Nova denominação para o complexo matogrossensis)
(Hemiptera, Reduviidae, Triatominae) do Estado do Rio Grande do Sul, Brasil.
Mem Inst Oswaldo Cruz 96: 71-79. [ Medline
] [ Lilacs
] [ SciELO
]
- Cortéz MR, Gonçalves
TCM 1998. Resistance to starvation of Triatoma rubrofasciata (De Geer,
1773) under labotratory conditions (Hemiptera: Reduviidae: Triato-minae).
Mem Inst Oswaldo Cruz 93: 549-554. [ Medline
] [ Lilacs
] [ SciELO
]
- Dias JCP 1965. Observações
sobre o comportamento de triatomíneos brasileiros frente ao jejum,
em laboratório. Rev Bras Malariol D Trop 17: 55-63.
[ Medline
]
- Dias-Lima AG, Sherlock IA 2001.
Períodos de resistência em jejum de algumas espécies de
triatomíneos (Hemiptera: Reduviidae: Triatominae). Rev Soc
Med Trop 34 (Supl. I): 140.
- Dias JCP, Feitosa VR, Ferraz-Filho
AN, Rodrigues VLC, Alencar SA, Sessa PA 1989. Fonte alimentar potencial vetorial
de Triatoma vitticeps (Stal, 1859) com relação à
doença de Chagas humana no Estado do Espírito Santo, Brasil.
Mem Inst Oswaldo Cruz 84 (Suppl. IV): 165-173.
- Ferreira E, Souza PA de, Fonseca
Filho M da, Rocha I 1986. Nota sobre a distribuição geográfica
do Triatoma vitticeps (Stal, 1859) (Hemiptera, Reduviidae) no Estado
do Rio de Janeiro, Brasil. Rev Bras Malariol D Trop 38: 11-14.
[ Medline
]
- Galvão C, Jurberg J, Cunha
V, Mello RP 1995. Biologia do Triatoma nitida Usinger, 1939 em laboratório
(Hemiptera: Reduviidae). Mem Inst Oswaldo Cruz 90: 657-663.
[ Medline
] [ Lilacs
]
- Galvão C, Jurberg J, Lent
H 1996. Resistência ao jejum de Triatoma nítida Usinger,
1939 em laboratório (Hemiptera: Reduviidae). Mem Inst Oswaldo Cruz
91: 639-640. [ Medline
] [ Lilacs
]
- Gonçalves TCM, Cunha V,
Oliveira E, Jurberg J 1997. Alguns aspectos da biologia de Triatoma pseudomaculata
Corrêa & Espínola 1964, em condições de
laboratório (Hemiptera: Reduviidae: Triatominae). Mem Inst Oswaldo
Cruz 92:275- 280. [ Medline
] [ Lilacs
] [ SciELO
]
- Gonçalves TCM, Oliveira
E, Dias LS, D Almeida M, Nogueira WO, Pires FDA 1998. An investigation on
the ecology of Triatoma vitticeps (Stal, 1859) and its possible role
in the transmission of Trypanosoma cruzi, in the locality of Triunfo,
Santa Maria Madalena municipal district, State of Rio de Janeiro, Brazil.
Mem Inst Oswaldo Cruz 93: 711-717. [ Medline
] [ Lilacs
] [ SciELO
]
- Gonçalves TCM, Victório
VNM, Jurberg J, Cunha V. 1988. Biologia do Triatoma vitticeps (Stal,
1859) em condições de laboratório (Hemiptera, Reduviidae,
Triatominae) I. Ciclo evolutivo. Mem Inst Oswaldo Cruz 83:519-523.
[ Medline
] [ Lilacs
]
- Gonçalves TCM, Victório
VMN, Jurberg J, Cunha V 1989. Biologia do Triatoma vitticeps (Stal,
1859) em condições de laboratório (Hemiptera, Reduviidae,
Triatominae). II - Resistência ao jejum. Mem Inst Oswaldo Cruz 84:
131-134.
- Heitzmann-Fontenelle T 1972. Bionomia
de Triatoma pseudomaculata Correa & Espínola, 1964, em laboratório.
Mem Inst Butantan 36: 251-262.
- Lima MM, Rey L, Mello RP de 1991.
Lethality of triatomines (Hemiptera: Reduviidae), vectors of Chagas' disease,
feeding on blood baits containing synthetic insecticides, under laboratory
conditions. Rev Inst Med Trop São Paulo 34: 295-301.
- Martinez-Ibarra J A, Katthain-Duchateau
G 1999. Biology of Triatoma pallidipenis (Stal, 1872) (Hemiptera: Reduviidae:
Triatominae ) under laboratory conditions. Mem Inst Oswaldo Cruz 94:
837-840.
- Moreira CJC, Perlowagora-Szumlewicz
A 1997. Attempts to improve xenodiagnosis: comparative test of sensibility
using Rhodnius neglectus, Panstrongylus megistus, Triatoma vitticeps
and Triatoma infestans in endemic areas of Brazil. Mem Inst Oswaldo
Cruz 92: 91-96. [ Medline
] [ Lilacs
] [ SciELO
]
- Neiva A, Lent H 1936. Notas e
comentários sobre triatomíneos. Lista de espécies e sua
distribuição geográfica. Rev Ent Rio de Janeiro 6:
153-190.
- Peçanha PM, Sessa PA, Silva
FJD, Pereira RS, Almeida IM 1983. Apresentação de três
casos autóctones do Estado do Espírito Santo. XIX Congresso
da Soc Bras Med Trop, Rio de Janeiro, p. 33.
- Pellegrino J 1952. Observações
sobre a resistência do Triatoma infestans ao jejum. Rev Bras
Biol 12: 317-320.
- Perlowagora-Szumlewicz A 1969.
Estudos sobre a biologia do Triatoma infestans, o principal vetor da
doença de Chagas no Brasil (importância de algumas de suas características
biológicas no planejamento de esquemas de combate a esse vetor). Rev
Bras Malariol D Trop 21: 117-159.
- Perlowagora-Szumlewicz A, Muller
CA, Moreira CJC 1990. Studies in search of a suitable experimental insect
model for xenodiagnosis of hosts with Chagas' disease. 4 - The reflection
of parasites stock in the responsiveness of different vector species to chronic
infection with different Trypanosoma cruzi stocks. Rev Saúde
Públ São Paulo 24: 165-177. [ Medline
] [ Lilacs
]
- Pinto AFS, Santos UM, Murad V,
Lima FEP, Zaganelli FL, Almeida AZ 1969. Doença de Chagas no Estado
do Espírito Santo. V - Primeiro caso de Chagas diagnosticado no município
de Alfredo Chaves, E.S. Rev Soc Bras Med Trop 3: 53.
- Pinto AFS, Vieira NER, Milanez
MC, Lima Pereira FE, Sessa PA 1986. Doença de Chagas autóctone
no Espírito Santo: relato de um caso agudo em gestante com transmissão
para o feto. Rev Soc Bras Med Trop 19 (Supl.): 78.
- Santos UM, Pinto AFS, Zaganelli
FL, Almeida AZ, Martins A, Carancho PV, Vieira HJ 1969a. Doença de
Chagas no esta-do do Espírito Santo. II - Inquérito sorológico.
E.S. Rev Soc Bras Med Trop 3: 51.
- Santos UM, Pinto AFS, Zaganelli
FL, Almeida AZ, Martins A, Carancho PV, Vieira HJ 1969b. Doença de
Chagas no estado do Espírito Santo. III - Vetores do tripanosoma. Rev
Soc Bras Med Trop 3: 51-52.
- Santos UM, Pinto AFS, Zaganelli
FL, Almeida AZ, Martins A, Carancho PV, Vieira HJ 1969c. Doença de
Chagas no estado do Espírito Santo. IV - Primeiro caso de Chagas diagnosticado
no município de Rio Novo do Sul, E.S. Rev Soc Bras Med Trop 3:
52-53.
- Sessa PA, Carias VRD 1986. Infecção
natural em triatomíneos do Espírito Santo por flagelados morfologicamente
semelhantes ao Trypanosoma cruzi. Rev Soc Bras Med Trop 19:
99-100. [ Medline
] [ Lilacs
]
- Silva CS, Santos-Mallet JR, Ziccardi
M, Gonçalves TCM 2001. Caracterização morfológica
e ultraestrutural de amostras de Trypanosoma cruzi, isoladas de Triatoma
vitticeps no estado do Rio de Janeiro. Rev Soc Med Trop 34 (Supl.
I): 122.
- Silva IG 1985. Influência
da Temperatura na Biologia de 18 Espécies de Triatomíneos (Hemiptera:
Reduviidae) e no Xenodiagnóstico, MSc Thesis, Universidade Federal
do Paraná, Curitiba, 169 pp.
- Silveira AC, Alencar TA, Máximo
MHC 1983. Sobre o Triatoma vitticeps Stal, 1859 (Hemiptera: Reduviidae)
no Espírito Santo, Brasil. X Reunião Anual de Pesquisa Básica
em Doença de Chagas, 15.
- Silveira AC, Feitosa VR, Borges
R 1984. Distribuição de triatomíneos capturados no ambiente
domiciliar no período de 1975-1983, Brasil. Rev Bras Malariol
D Trop 36: 15-312. [ Medline
]
- Uribe C 1926. On the biology and
the life history of Rhodnius prolixus Stal, 1859. J Parasitol 13:
129-136.
Copyright 2002 Instituto Oswaldo
Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc02232t1.jpg]
[oc02232t5.jpg]
[oc02232t3.jpg]
[oc02232t2.jpg]
[oc02232t4.jpg]
[oc02232f1.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}