Mem Inst Oswaldo Cruz, Rio de
Janeiro, Vol. 97(7), October
2002, pp. 1063-1065
Feeding and Defaecation
Behaviour of Triatoma patagonica (Del Ponte, 1929) (Hemiptera: Reduviidae)
Julieta Nattero+,
Liliana B Crocco, Claudia S Rodríguez
Cátedra de Introducción
a la Biología, Facultad de Ciencias Exactas Físicas y Naturales,
Universidad Nacional de Córdoba, Vélez Sarsfield 299, 5000 Córdoba,
Argentina
+Corresponding author. Fax: +54-351-433.1056 E-mail:
jnattero@efn.uncor.edu
This work received
financial support from Secyt (Argentina).
Received 10 January 2002
Accepted 22 July 2002
Code Number: oc02234
Among the vectors of Chagas disease,
Triatoma patagonica is a species in the process of adaptation to the
human environment being recently registered in urban and suburban zones. However,
its importance as a vector of Chagas disease is unknown. The aim of this work
was to evaluate two aspects of vectorial competence: the feeding behaviour and
the defaecation pattern. These processes were studied in females of T. patagonica
fed ad libitum on a restrained pigeon. The results showed that the blood meal
size was negatively correlated with the time of first defaecation (r = -0.42).
The first defaecation was emitted before the first 10 min and defaecations during
feeding were frequent. A total of 73% of females, defaecated during the first
30 min post-feeding. These results suggest that if this species subsequently
colonizes the domicile, it would be capable of transmitting Trypanosoma
cruzi.
Key words: Triatoma patagonica
- feeding - defecation - peridomestic - Chagas disease vector - Argentina
Among the peridomestic species of
Chagas disease vectors, Triatoma patagonica (Del Ponte, 1929) has the
most southerly distribution. It is only to be found in Argentina (Lent &
Wygodzinsky 1979), 20º to 50º southern latitude and 750 m of altitude
(Carcavallo et al. 1999).
It was found naturally infected by
Trypanosoma cruzi (Chagas 1909) (Lent & Wygodzinsky 1979) and in
Río Colorado (province of Río Negro) it was the only peridomestic
species found in houses both in urban and suburban areas (Ferrero et al. 1999).
These last records, added to the permanence in the peridomicile, point the need
to determine how competent T. patagonica is as a vector of Chagas disease,
especially in aspects related to feeding and defaecation.
These two processes (feeding and
defaecation) have been thoroughly analyzed in triatomines as they are closely
related to the development and transmission of T. cruzi (Mello 1980,
Kirk & Schofield 1987, Trumper & Gorla 1991, Crocco & Catalá
1996). These processes are well known in domestic species like T. infestans
(Klug, 1934) (Forattini et al. 1982, Friend & Smith 1985, Trumper &
Gorla 1991) and Rhodnius prolixus (Stal, 1859)(Forattini et al. 1982,
Friend & Smith 1985). Nevertheless, very little is known about peridomestic
species including T. patagonica (Del Ponte, 1929) about which only facts
relating to its life cycle and habitats are known (Martínez et al. 1985,
Carcavallo et al. 1998).
From an epidemiological point of
view, it is very important to have knowledge related to feeding and defaecation,
the aim of this work is to evaluate the feeding pattern and the defaecation
behaviour in adults of T. patagonica.
MATERIALS AND METHODS
This work was carried out with 29
adult females of T. patagonica. These were obtained as fifth instar nymphs
by the Servicio Nacional de Chagas (Argentina). The insects were maintained
under laboratory conditions at 26ºC ± 1ºC and at 60-70% humidity.
After moulting, the insects were weighed and marked with acryllic paint following
predetermined codes and were starved for 15 days (after moulting). After fasting,
they were fed ad libitum on a restrained pigeon. For each feeding the following
variables were determinated:
Blood meal size - This was
determined by the difference in weight before and after feeding.
Feeding time - This was obtained
by recording the time (in minutes) since the rostrum of the insect came into
contact with the skin of the host until it detached itself.
Number of defaecations during
feeding - This was registered by direct observation of the insect while
feeding on the pigeon.
Number of defaecations emitted
during the first 30 min after feeding - This was registered by direct observation
of the insect while it was on the host and during 30 min after feeding.
All weights were measured using a
Mettler Balance with a precision of 0.001 mg. The t-test for samples with dependent
variables for comparing variables of the different groups, was used as statistic
treatment. To verify the homogeneity of variance the Levenne test was used (Ferrán
Aranaz 2001).
RESULTS
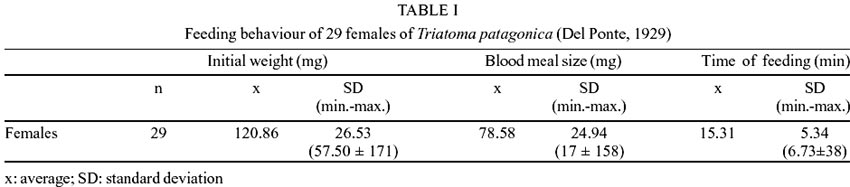
In Table
I, the variables analyzed concerning feeding behaviour of females of T.
patagonica are shown. The data were obtained from the repletion feeding
of 29 females of T. patagonica, the average time recorded to complete
an ad libitum ingest was approximately 15 min (SD = 5.34) although this time
was not related to the blood meal size, which varied between 17 to 158 mg.
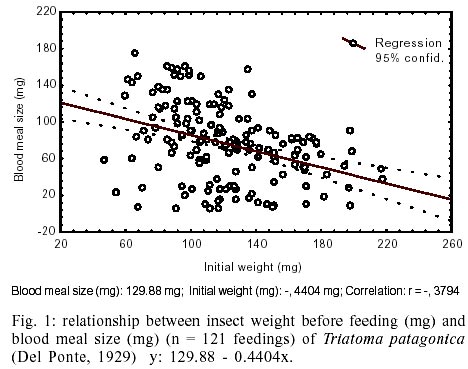
In order to establish if the nutritional
state of the insect (weight before each feeding) affected the size of each blood
meal ingestion, the analysis of correlation between the initial weight (mg)
and the blood meal size (mg) was carried out (Fig.
1). These analysis showed a negative linear correlation (r = -0.38, n =
172, p < 0.0500), with higher weight insects ingesting less, for example
on insects with an initial weight of 57.5 mg (minimum registered) will take
a medium ingest of 104.56 mg.
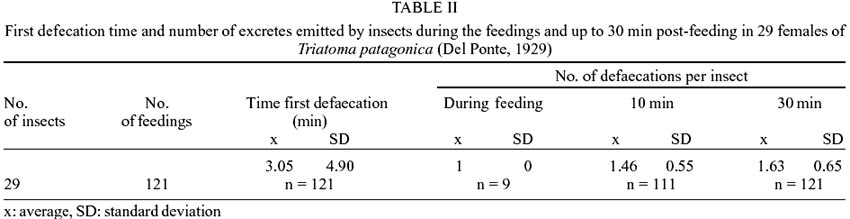
On the other hand the defaecating
behaviour was analysed from 158 ingest ad libitum. These feedings began 15 days
after moulting to adults and during all the insect life. It was observed that
from the total number of feedings, there were excretions in a 73% of the cases
(n = 121). Significant differences in the size of the ingest were registered
between females that defaecated (x = 89.86 mg) and those that did not defaecate
(x = 45.26 mg) (p < 0.0500).
In terms of the number of defaecations,
an average number was obtained of 1.00, 1.46 and 1.63 defaecations per insect
during the feeding and at 10 and 30 min post ingestion, and at 10 min they were
registered up to 3 defaecations. The average time for the first defaecation
was at 3.05 min ± 4.90 and the second one occurred within 10 min (Table
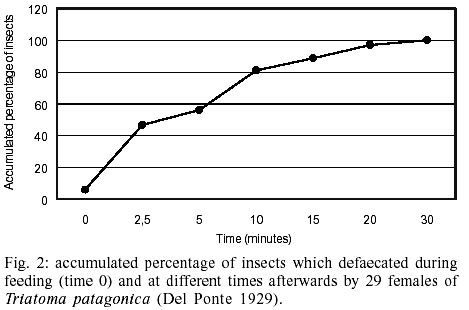
II). The accumulated percentage of defaecations per insects was analyzed
at different time. For the total of insects that defaecated during the first
30 min post ingest. In Fig. 2 it
can be observed that at 2.5 min post ingestion, close to 50% of the females
had emitted excretions and at 10 min post ingest the value recorded was 80%.
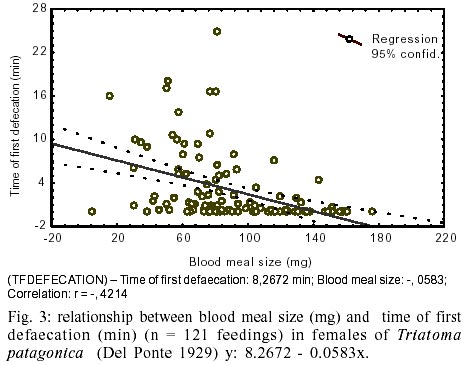
In order to determinate if the time
of the first defaecation depends on the size of the ingestion (blood meal size)
a correlation analyzed using both variables was carried out. A negative correlation
was found (r = -0.42, n = 121, p < 0.0500), the larger the ingest the shorter
the time for the first excretion. For an ingest of 60 mg, the time recorded
for the first excretion was 4.8 min (y: 8.2672- 0.0583.x) (Fig.
3).
DISCUSSION
The results obtained show that females
T. patagonica require approximately 15 min to feed ad libitum. The size
of this ingest was variable (x = 78.19 mg) but in general did not exceed the
insect weight.
A comparison with T. infestans
and the peridomestic species T. sordida (Stal, 1859) using the index
of relative ingest (IIR), revels that the capacity of T. patagonica
(0.6) is smaller than that of T. infestans (1.1) (Perlowagora 1973) and
T. sordida (1.3) (Crocco & Catalá 1996).
On the other hand it can be observed
that defaecating behaviour depends on the size of the ingest. The larger the
ingest, the shorter the time between the begin of the feeding and the first
defaecation. The minimum ingest required to induce defaecation during the first
10 min was approximately 20 mg. This relationship between the defaecation time
and the size of the ingest were also registered in T. infestans (Trumper
& Gorla 1991) and in T. sordida (Crocco & Catalá 1996).
In triatomines when the ingestions are abundant, they must to emit the excretions
quickly, because the blood have too much water (Friend & Smith 1985).
T. patagonica, not only defecaeted quickly on the host after ingestion,
some insects defaecated during the feeding time. To defecaete during the feeding
time, T. patagonica needs a minimum ingest of 130 mg, more than that
required by T. sordida (80 mg).
When compared the percentage of insects
that defecaete at 10 min after feeding, the time considered by Zeledón
(1975) to offer the highest risk for the transmission of T. cruzi, it
can observed that T. patagonica presents a similar pattern to those shown
by T. infestans and T. sordida. At 10 min after feeding the percentage
of females of T. patagonica that defaecaeted was approximately 80% while
for T. infestans it was 92% (Zeledón et al. 1977) and for T.
sordida approximately 97% (Crocco & Catalá 1996).
It was registered that in T. patagonica
the nutritional state of the insect in the moment of feeding affects the blood
meal size. In T. sordida, studies carried out by Schofield et al. (1991),
demonstrated that the nutritional state of the insect influences the dispersion
of the species, given that the proportion of adults that start to fly is lower
when the nutritional state is higher. If we translate these data to T. patagonica
it suggests that insects with a low nutritional state will have more capacity
to disperse, making it possible to colonize new ecotopes. Moreover, insect with
a lower nutritional state ingest, a greater quantity of blood increasing the
possibility of defecation during the feeding.
Finally, it can be concluded that
the result obtained here suggest that if T. patagonica werereally to
colonize the domiciles, it would be capable of transmission of T. cruzi,
because it is a species that has the characteristic of defaecating immediately
after as well as during feeding.
ACKNOWLEDGMENT
To the Servicio Nacional de Chagas
for supplying insects.
REFERENCES


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}