 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(8), December 2002, pp. 1111-1116 Effect of Sequential Heat and Cold Shocks on Nuclear Phenotypes of the Blood-sucking Insect, Panstrongylus megistus (Burmeister) (Hemiptera, Reduviidae) Simone L Garcia*, Raquel M Pacheco**, Vera LCC Rodrigues***, Maria Luiza S Mello/+
Departamento de Biologia Celular
*Departamento de Parasitologia, Instituto de Biologia **Departamento de Estatística,
Instituto de Matemática, Estatística e Computação
Científica, Unicamp, 13084-971 Campinas, SP, Brasil ***Sucen, Mogi Guaçu,
SP, Brasil This study was supported by the State of São Paulo Research Foundation (Fapesp, grants no. 98/0826-4 and 99/2547-8) and the Brazilian National Research and Development Council (CNPq). Received 16 January 2002 Code Number: oc02243 Thermal shocks induce changes in the nuclear phenotypes that correspond to survival (heterochromatin decondensation, nuclear fusion) or death (apoptosis, necrosis) responses in the Malpighian tubules of Panstrongylus megistus. Since thermal tolerance increased survival and molting rate in this species following sequential shocks, we investigated whether changes in nuclear phenotypes accompanied the insect survival response to sequential thermal shocks. Fifth instar nymphs were subjected to a single heat (35 or 40°C, 1 h) or cold (5 or 0°C, 1 h) shock and then subjected to a second shock for 12 h at 40 or 0°C, respectively, after 8, 18, 24 and 72 h at 28°C (control temperature). As with specimen survival, sequential heat and cold shocks induced changes in frequency of the mentioned nuclear phenotypes although their patterns differed. The heat shock tolerance involved decrease in apoptosis simultaneous to increase in cell survival responses. Sequential cold shocks did not involve cell/nuclear fusion and even elicited increase in necrosis with advancing time after shocks. The temperatures of 40 and 0oC were more effective than the temperatures of 35 and 5oC in eliciting the heat and cold shock tolerances, respectively, as shown by cytological analysis of the nuclear phenotypes. It is concluded that different sequential thermal shocks can trigger different mechanisms of cellular protection against stress in P. megistus, favoring the insect to adapt to various ecotopes.
Key words: heat shock - cold shock - sequential shocks - nuclear phenotypes - Panstrongylus megistus Panstrongylus megistus, one of the most important vectors of Chagas disease, is a blood-sucking insect with a wide geographical distribution in Brazil, high rates of infection with Trypanosoma cruzi, and an adaptability for invading artificial ecotopes (Forattini 1980). Prolonged cold and heat shocks are deleterious to P. megistus nymphs (Garcia et al. 1999), but resistance to these stressing conditions can be elicited following sequential shocks in which a mild shock precedes a severe one (Garcia et al. 2001a, b). Heat-shock tolerance follows a pattern typical of heat shock protein activation (Garcia et al. 2001a). Cold-shock tolerance also presumably involves heat shock proteins, although consideration of other protective mechanisms has not been eliminated (Garcia et al. 2001b). The normal nuclear phenotype in P. megistus males has a small heterochromatic body which contains several copies of the Y chromosome (Mello et al. 1986), but phenotypes indicative of survival (decondensed heterochromatin, nuclear fusion) and death (apoptosis, necrosis) responses have also been found in male nymphs and adults. Nuclei with heterochromatin loosening may represent an attempt to activate silent genes during stress (Mello 1983, 1989). Giant nuclei are characterized by a bigger size when compared with normal nuclei, and originate from nuclear and/or cellular fusion (Wigglesworth 1967, Mello & Raymundo 1980). This phenotype has been described as a possible mechanism of cellular and organ protection under unfavourable conditions in blood-sucking insects (Wigglesworth 1967, Mello & Raymundo 1980, Dantas & Mello 1992, Garcia et al. 2000a, b). Apoptosis and necrosis are defined in terms of their morphological characteristics (Kerr 1971, Kerr et al. 1972, Garcia et al. 2000a, b). With the exception of the normal phenotype, the other phenotypes are more frequent in specimens subjected to heat and cold shocks (Garcia et al. 2000a, b). The frequency of fused nuclei and of nuclei with heterochromatin decondensation compared with that of other phenotypes was considered insufficient to cope with the deleterious effects of a severe heat shock (Garcia et al. 2000a). For the response to cold shock, the activation of nuclear and cell fusion which generates giant nuclei was found to be less effective or necessary than that elicited by heat shock (Garcia et al. 2000b). Considering the cell survival and cell death responses expressed by nuclear phenotype changes in P. megistus subjected to single heat and cold shocks (Garcia et al. 2000a, b) and the increase in specimen survival and molting rates in this species under sequential heat and cold shocks (Garcia et al. 2001a, b), we suspected that changes in nuclear phenotypes may also occur under sequential temperature shocks. In the present study, the influence of sequential heat and cold shocks on nuclear phenotypes was investigated in Malpighian tubule epithelial cells of P. megistus. MATERIALS AND METHODS
Insects and shock assays - Male fifth instar nymphs of P. megistus (Burmeister) (Hemiptera, Reduviidae) were reared at 28°C and 80% relative humidity and fasted for 15 days prior to the temperature shock experiments, since only with moderate fasting do some nymphs survive prolonged heat and cold shocks (Garcia et al. 1999). The insects were subjected to a shock at 35 or 40°C for 1 h, returned to 28°C and then subjected to a further shock at 40°C for 12 h. Other groups of insects were subjected to a shock at 5 or 0°C for 1 h, returned to 28°C and then subjected to a further shock at 0°C for 12 h. The choice of the shock temperatures was based on previous reports (Garcia et al. 2001a, b). For both the heat and cold shock assays, the second shock started 8, 18, 24 and 72 h after the first one. After the second shock, the insects were returned to 28°C and fed on hen blood once a week. Control specimens were maintained at 28°C, the temperature used to rear P. megistus in Sucen's laboratory since 1980, or subjected to a single shock at 0, 5, 35 and 40°C, for 1 h, and 0 or 40°C for 12 h. Organ preparations - Malpighian tubule preparations were obtained immediately after the shocks or 10 and 30 days later. Organs from at least three specimens were used for each experimental condition and the corresponding control. The whole organs were mounted on glass slides, immediately fixed in ethanol: acetic acid solution (3:1, v/v) for 1 min, rinsed in 70% ethanol for 5 min, and air dried at room temperature. The tubules were then subjected to the Feulgen reaction, with acid hydrolysis in 4 M HCl at 25°C for 65 min. The Feulgen-stained material was rinsed in sulfurous and distilled water, air dried, cleared in xylene and mounted in Canada balsam. Nuclear counting and statistics - A Nikon light microscope was used to count the total number of Malpighian tubule epithelial cell nuclei per specimen and to determine the frequency of the different nuclear phenotypes present. To assess the relationship between stress conditions and the various nuclear phenotypes, analysis of variance (ANOVA) with two factors (interval between shocks and number of days following shocks) was used, considering an unbalanced number of nuclei with specific phenotypes for different specimens (Kalbfleish & Prentice 1980). The same tests used for the heat shock results were applied to cold shock data, except that severe cold shocks (12 h) were not tested because in this case almost all insects died. RESULTS
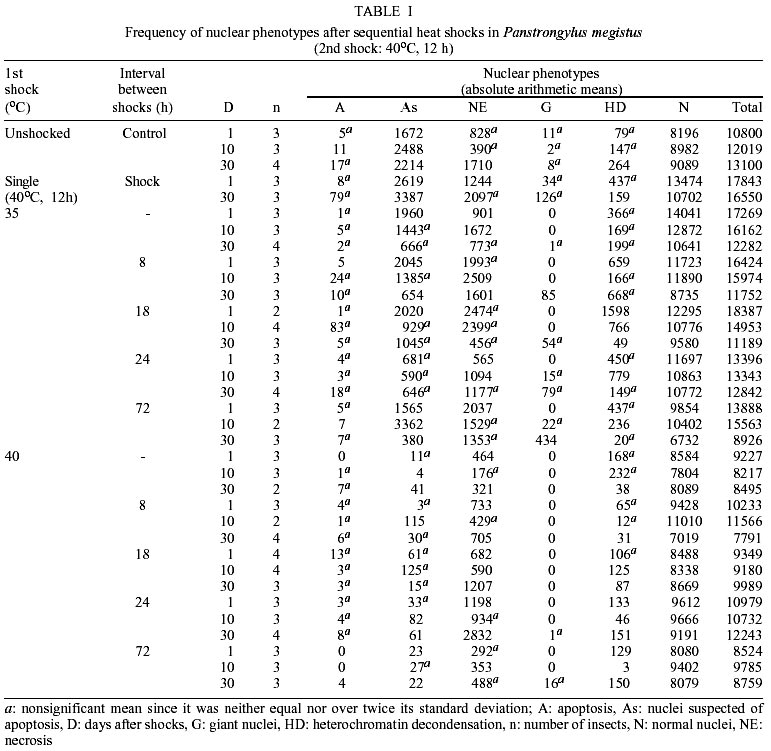
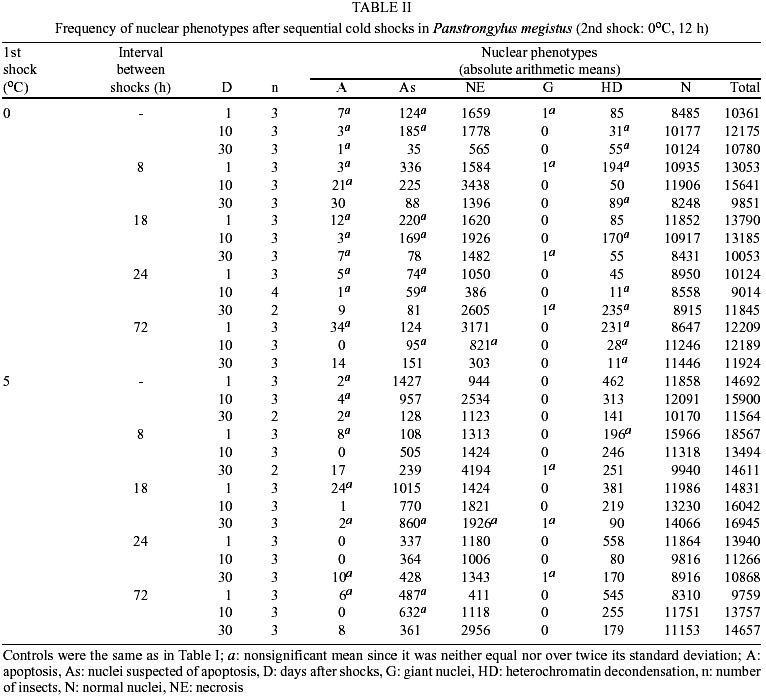
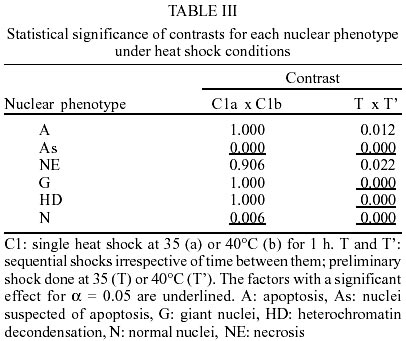
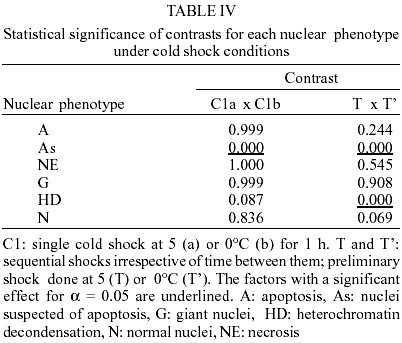
The same phenotypes observed in this work have been described and reported elsewhere (Mello 1978, Mello et al. 1986, Garcia et al. 2000a, b). The most frequent phenotype observed in the control groups and in the groups subjected to single or sequential shocks was the normal one. Among the changed phenotypes, there were nuclei with heterochromatin decondensation, as well as giant nuclei and nuclei with morphological aspects of typical and suspected apoptosis and necrosis. The frequencies of the nuclear phenotypes in the various experimental conditions are shown in Tables I and II. Apoptosis - The frequency of apoptosis in the insects that received sequential heat shocks did not differ from that of the unshocked group (p: 0.317), but did increase in comparison to that of the insects that received the single mild shock (p: 0.036) and did decrease in comparison to that of the specimens that received the single severe shock (p: 0.015). In general, with advancing time after the heat shock, the frequency of this phenotype increased (p: 0.004). When considering cold sequential shocks, the frequency of apoptosis decreased as compared with the unshocked group (p: 0.003), but did not differ from that of the specimens that received the single mild shock (p: 0.510). Change in shock temperature from 35 to 40oC or 5 to 0oC did not affect significantly the frequency of apoptosis in single heat and cold shock assays, respectively (Tables III, IV). However, it caused a decrease in frequency of this phenotype when a second heat shock was applied (Table III). Nuclei suspected of apoptosis - The frequency of this phenotype in the specimens subjected to sequential heat shocks decreased as compared with the unshocked control (p: 0.000) and the group that received the single severe shock (p: 0.000), but did not differ from that of the specimens that received the single mild shock (p: 0.791). Thirty days after the sequential shocks, the frequency of the nuclei suspected of apoptosis was lower than the average of the frequencies detected for 1 and 10 days after the shocks (p: 0.001). When the time between shocks was 24 h and the insects were analyzed 1 day after the shocks, the frequency of this phenotype was smaller than the average of the frequencies detected for the other periods between shocks (p: 0.001). When the time between shocks was 72 h and the insects were analyzed 10 days after shocks, the frequency of this phenotype increased (p: 0.004). Under cold shock conditions, the frequency of the nuclei suspected of apoptosis was generally similar to that verified after heat shocks. When a shock at 40 or 0oC was applied, irrespective of a second shock being given, there was a decrease in the frequency of nuclei suspected of apoptosis in comparison with shocks at 35 or 5oC, respectively (Tables III, IV). Necrosis - In general, sequential heat shocks did not affect the frequency of necrosis when compared with the unshocked group (p: 0.091) and the groups subjected to the single mild (p: 0.073) or severe (p: 0.904) shocks. When the period between shocks was equal to 18 h and the insects were dissected 30 days after the shocks, the frequency of necrosis was lower than the average of the frequencies obtained for longer periods between shocks (p: 0.002). No change in the frequency of necrosis was verified 1 day after sequential cold shocks. However, with advancing time after shock, the frequency of this phenotype increased significantly. Change in shock temperature from 35 to 40oC or 5 to 0oC did not affect the frequency of necrosis in single heat and cold shock assays, respectively (Tables III, IV). However, it promoted a decrease in frequency of necrosis when a second heat shock was given (Table III). Giant nuclei - Sequential shocks increased the frequency of giant nuclei in comparison with the group subjected to the single severe shock (p: 0.001), but did not differ from that of the insects that received the single mild shock (p: 0.612). The frequency of giant nuclei 30 days after heat shocks was greater than for the other times after shocks. In addition, sequential shocks decreased the frequency of giant nuclei relative to the unshocked group 1 day after shocks (p: 0.005), but increased the frequency of this phenotype 30 days after shocks (p: 0.002). When considering the application of cold shocks, the occurrence of giant nuclei decreased in comparison with the unshocked group (p: 0.000). Increasing the temperature of the preliminary heat shock from 35 to 40oC decreased the frequency of giant nuclei in sequential shock assays (Table III). No change was promoted in the frequency of this phenotype when the temperature of the preliminary shock in sequential cold shock assays increased from 0 to 5oC (Table IV). Heterochromatin decondensation - The frequency of this phenotype increased after sequential heat shocks in comparison with the unshocked group (p: 0.027) or with the groups subjected to the single mild or severe shock (p: 0.030 and 0.011, respectively). Under cold shock conditions, heterochromatin decondensation was more effective at short periods after sequential shocks. Change in shock temperature from 35 to 40oC or from 5 to 0oC had no effect on the frequency of heterochromatin decondensation in single heat and cold shock assays, respectively (Tables III, IV). However, it promoted a decrease in frequency of this phenotype when sequential shocks were given (Tables III, IV). Normal nuclear phenotype - The frequency of this phenotype did not differ when heat (p: 0.179) or cold (p: 0.187) sequential shocks were compared to the unshocked group, but decreased relative to that of insects subjected to the single mild shock (p: 0.018 - heat shock, p: 0.011 - cold shock). On the other hand, the frequency of normal nuclei in the groups subjected to sequential shocks was larger than in the group subjected to a severe shock only ( p: 0.036). Change in frequency of normal nuclei with the temperature of the preliminary or the single shock was only verified in the heat shock assay (Table III). The frequency of normal nuclei was larger when the temperature of the preliminary heat shock was 40oC. DISCUSSION
Nuclei with normal phenotype as well as with phenotypes indicative of cell survival (nuclear fusion and heterochromatin decondensation) and cell death (apoptosis and necrosis) were found concomitantly in the Malpighian tubules of P. megistus under most of the experimental conditions used. These phenotypes had been previously reported not only after single heat and cold shocks in P. megistus (Garcia et al. 2000a, b), but also after single heat and cold shocks and sequential cold shocks in Triatoma infestans (Dantas & Mello 1992, Mello et al. 2001, Campos et al. 2002). Heat shock tolerance has been demonstrated for P. megistus in terms of increased specimen survival and molting rates elicited by sequential heat shocks in comparison with single heat shocks (Garcia et al. 2001b). Present results indicate that heat shock tolerance in P. megistus also involves morphological changes at the cellular level. The expression of these changes is characterized by decrease in a cell death form (apoptosis) simultaneously to an increase in cell survival responses (Garcia et al. 2000b, Mello et al. 2001), especially when comparisons are made with results obtained after a single severe shock. The heat shock tolerance in terms of protecting cell mechanisms elicited by sequential heat shocks being more efficient and longer lasting was verified when the temperature of the 1st shock was 40oC in comparison with the shock at 35oC. A more drastic change in temperature from 28 (control) to 40oC is possibly required to activate massive alterations in genomic expression, with numerous specialized metabolic responses thus being triggered (Gasch et al. 2000). Cold shock tolerance, also demonstrated in P. megistus in terms of increased specimen survival and molting rates after sequential cold shock assays (Garcia et al. 2001a), was equally accompanied by changes in frequency of specific cell nuclear phenotypes. However, the frequency of these changes were not always the same as those observed after sequential heat shocks. Apoptosis, for instance, decreased significantly after sequential cold shocks only in comparison with unshocked controls. While cell/nuclear fusion increased with sequential heat shocks, this form of cell survival (Wigglesworth 1967, Mello 1989, Mello et al. 2001) decreased with sequential cold shocks. As regards the cellular response to sequential cold shocks when the temperature of the 1st shock was 0oC in comparison to 5oC, it differed from the situation considered in the sequential heat shock assays. A more drastic change in temperature from 28 to 0oC revealed a reduced ability or need of the cells to cope with this condition when examining the decrease in frequency of heterochromatin decondensation in relation to that promoted when the 1st shock was given at 5oC. However, this difference in frequency of nuclear phenotypes did not affect the insect survival rate (Garcia et al. 2001a). The frequency of necrosis was differently affected when heat and cold shock assays were compared to each other. It increased significantly under sequential cold shock conditions. As regards the response to this experimental condition, the same agents which induce decrease in frequency of apoptosis may be responsible for increase in frequency of necrosis with advancing time after shocks. Among these factors, change in the cellular ATP/AMP ratio promoted by the stress might be causing a sudden decrease in cellular ATP concentrations (Hardie 1999, Gasch et al. 2000) thus favoring metabolic conditions to death through necrosis. In conclusion, as with specimen survival (Garcia et al. 2001a,b), different patterns of changes in the nuclear phenotypes were induced by sequential heat and cold shocks in P. megistus. These results demonstrate that different sequential thermal shocks can elicit different mechanisms of cellular protection against stress in this insect. This is in agreement with observations in other cell models showing that although a large set of genes (~ 900 in yeast cells) shows a similar response to almost all environmental changes, additional features of the genomic responses are specialized for specific conditions (Gasch et al. 2000). This variable capacity may allow P. megistus to adapt to various ecotopes over a wide geographic range
ACKNOWLEDGMENTS
To Dr Stephen Hyslop for revision of the manuscript and to Mr Pedro Ribeiro da Silva for technical help. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02243t1.jpg] [oc02243t2.jpg] [oc02243t3.jpg] [oc02243t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}