 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(8), December 2002, pp. 1153-1156 Detection of hilA Gene Sequences in Serovars of Salmonella enterica Subspecies Enterica Nora Cardona-Castro/+, Eliana Restrepo-Pineda, Margarita Correa-Ochoa*
Laboratorio de Microbiología,
Instituto Colombiano de Medicina Tropical, Cra 43ª No. 52 Sur 99. AA 52162,
Sabaneta, Antioquia, Colombia *Escuela de Bacteriología, Universidad
de Antioquia, Medellín, Colombia This project was supported by grant 3256-04-304-98 from Colciencias. Received 8 February 2002
Code Number: oc02251 hilA gene promoter, component of the Salmonella Pathogenicity Island 1, has been found in Salmonella serovar Typhimurium, being important for the regulation of type III secretion apparatus genes. We detected hilA gene sequences in Salmonella serovars Typhi, Enteritidis, Choleraesuis, Paratyphi A and B, and Pullorum, by polymerase chain reaction (PCR) and hybridization techniques. The primers to carry out PCR were designed according to hilA sequence. A low stringency hybridization with the probe pVV441 (hilA open-reading-frame plasmid) was carried out. To find hilA gene sequences in other Salmonella sp. suggest that these serovars could have similar sequences of this kind of virulence genes.
Key words: hilA gene sequences - Salmonella serovars Typhi - Enteritidis - Choleraesuis - Paratyphi A and B - Pullorum The pathogenic mechanisms of Salmonella enterica serovar Typhimurium have been the most widely studied among Salmonella species, since it is easily manipulated with molecular biology techniques, as well as for having an animal model that allows in vivo experiments (Lee 1994). Several genes have been identified and associate with the complex mechanisms that S. enterica serovar Typhimurium uses to invade epithelial intestinal cells (Darwin & Miller 1999, 2001, Lostroh et al. 2000, Detweiler et al. 2001, Lucas & Lee 2001). Salmonella Pathogenicity Island 1 (SPI1) is composed by several genes, hilA (hyperinvasive locus A), is one of them (Lee 1994, 1996, Lee et al. 1992, Bajaj et al. 1995, Lostroh et al. 2000, Milrod et al. 2001) and it is required for the regulation of the type III secretion genes (Lostroh et al 2000, Lesnick et al. 2001). Virulence genes like type III secretion apparatus, secret proteins that are probably related to cell invasion (Kubori et al. 1998, Darwin & Miller 2001, Lesnick et al. 2001). S. enterica serovar Typhimurium produces infection in the host and penetrates the epithelial cells in the membrane-associated vacuoles (Kubori et al. 1998, Lostroh et al. 2000); also, can induce apoptosis (Zhou et al. 2000, Santos et al. 2001), kill macrophages by a caspase-1-dependent mechanism that also releases proinflammatory cytokines. Caspase-1 activation correlates with the ability to colonize the lymph nodes, spleen and liver in mice (Detweiler et al. 2001). All of the mechanisms described allow S. enterica serovar Typhimurium to cross the intestinal epithelial barrier and cause systemic infection (Miras et al. 1995, Kubori et al. 1998, Lostroh et al. 2000, Zhou et al. 2000, Detweiler et al. 2001, Santos et al. 2001). In mice, S. enterica serovar Typhimurium replicates in macrophages; it is thought that either macrophages or dendritic cells carry the bacteria from the Peyer's patches to the lymph nodes, spleen and liver and produces a systemic, typhoid-like disease (Miller et al. 1995, Darwin & Miller 1999, Detweiler et al. 2001). In immunocompetent humans, S. enterica serovar Typhimurium does not cause systemic disease like serovars Typhi and Paratyphi A, B, C, and bacterial replication is limited to the intestine producing gastroenteritis. It is unclear why S. enterica serovar Typhimurium causes different diseases in mice and humans and why S. enterica serovar Typhi can produce systemic infections in humans and other serovars can not do it (Detweiler et al. 2001, Fierer & Guiney 2001). Other Salmonella sp. produces in humans several clinical syndromes such as, typhoid fever by serovar Typhi and Paratyphi A, B, C, gastroenteritis by Typhimurium, Newport, Enteritidis and Pullorum exceptionally; bacteremia by serovar Choleraesuis, as well as localized tissue infections (Miller et al. 1995), the pathogenic mechanisms of these Salmonella serovars have not been studied. To search pathogenic mechanisms in non Typhimurium Salmonella sp., should contribute to understand different clinical syndromes, that Salmonella sp. produce in humans. We searched and detected hilA gene in clinical important serovars of S. enterica subsp. enterica: Typhi, Paratyphi A and B, Pullorum, Choleraesuis, and Enteritidis. MATERIALS AND METHODS S. enterica serovar Typhimurium SL 1344 was used as hilA control strain. Plasmid pVV441, which is a Bluescript II KS+ (Stratagene, USA) derivative that carries a 1.6 Kb BamHI fragment generated by polymerase chain reaction (PCR) and, except for the first 60 nucleotides and last 20 nucleotides, contains the hilA open-reading-frame (communication from Dr Catherine A Lee), was used as DNA probe for low stringency hybridization test and positive control for PCR. S. enterica serovar Typhi 404Ty is a DaroAl48 derivative of an Indonesian strain, 3083/30, which has two flagellar antigen phases, d and z66 (Lee & Falkow 1990), Enteritidis 91A 3404, Pullorum 159 and Choleraesuis SL 2824, were used to search hilA gene in their chromosomes by hybridization and PCR tests. DNA from clinical isolates of S. enterica serovar Typhi, Enteritidis, Pullorum, Choleraesuis and Paratyphi A and B, Vibrio cholerae O1, Proteus mirabilis, Klebsiella pneumoniae, Pseudomonas aeruginosa, Citrobacter freundii, Escherichia coli and Shigella flexneri, were tested for hilA gene by PCR. Bacterial DNA extraction was carried out according the methodology described by Graves and Swaminathan (1995), using lysozyme (Sigma Chemical Co, St Louis, MO, USA) and phenol chloroform extractions. Chromosomal DNA was digested using EcoRI and EcoRI plus BamHI, SalI and SalI plus HindIII (New England Bio Labs, Inc. Beverly MA, USA). A Southern blot and low stringency hybridization test using DNA probe pVV441 was carried out (Swaminathan & Matar 1995, Tenover & Unger 1995). The probe was labeled and detected by chemiluminescence by ECL system following the manufacturer recommendations (Amersham International, Amersham, UK). Primers to carry out PCR were synthesized according to hilA sequences (854 bp) at gene library (GenBank-today U25352, www.genome.ad.jp). The sequences are: DS 5'-CGG AAG CTT ATT TGC GCC ATG CTG AGG TAG-3' and US 5'-GCA TGG ATC CCC GCC GGC GAG ATT GTG-3' PCR was performed in a Programmable Thermal Controler (PTC-100, MJ Research, Inc., Watertown, MA) using 50 µl reaction volume containing 100 mM Tris-HCl (pH 9.0), 500 mM KCl, 1.0% Triton X-100, 25 mM MgCl2, 0.20 mM of each deoxynucleoside triphosphate (Promega, Madison, WI), 20 µM of each primer, 0.5 U of Taq polymerase (Promega) and 3 µl of bacterial DNA. The parameters for amplification were as follows: initial denaturation at 94°C for 3 min, 30 cycles of 94°C for 1 min, 65°C for 1 min, 72°C for 1 min and a final extension step at 72ºC for 10 min. The PCR products were electrophoresed in a 2% agarose gel. After staining with ethidium bromide, the amplified DNA fragment in the gel was visualized. The molecular weight marker used was f X174 DNA/Hae III Markers (Promega). RESULTS
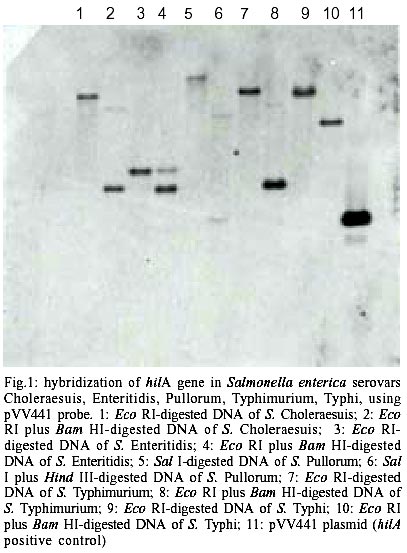
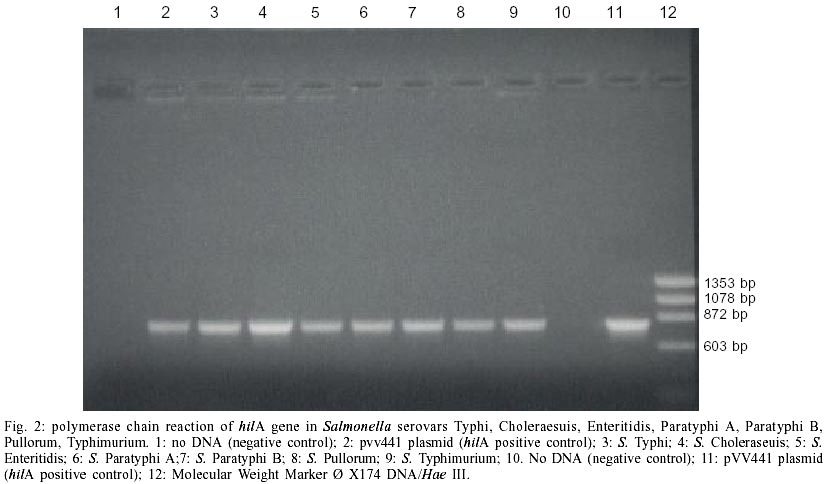
Figs 1 and 2 show the hilA gene found by hybridization and PCR respectively. A 854 bp band detected by PCR, was considered like hilA gene, this correspond to the size of the gene (GenBank-today U25352, www.genome.ad.jp). The results of hybridization were comparable to PCR results, both techniques were consistent in finding the hilA gene in all Salmonella sp. PCR to detect hilA carried out in bacteria different to Salmonella was negative in all cases. Detection of hilA gene sequences in other Salmonella serovars that cause human infections, is new and it tells about the presence of the same gene that was found in S. enterica serovar Typhimurium involves in the regulation of the type III secretion genes. The virulence genes type III are important because they coded the secretion of proteins related with cell invasion. Guo et al. (2000), used PCR to detect hilA gene in tomatoes contaminated with S. enterica serovar Montevideo; this report suggests that hilA gene is present in other serovar that we did not test, increasing the importance of this data. Until now, the Salmonella pathogenesis was focused in S. enterica serovar Typhimurium and the results were transferred to other Salmonella sp. Popoff's group found in S. enterica serovar Typhi an almost identical sequence that hilA gene and called it iagA (Miras et al. 1995). The utilization of two different methodologies, PCR and hybridization, that detected hilA gene sequences make more confident these results that can be used in clinical, industrial and research fields. However to amplify a sequence or detect it by hybridization does not mean that it is functional, it is necessary to test the function of hilA gene in these Salmonella species.
ACKNOWLEDGMENTS
To Dr Catherine A Lee for tutorial support, and for providing Salmonella strains and hilA gene probes. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02251f2.jpg] [oc02251f1.jpg] |
| |||||||||

{kind=link}
{kind=link}