 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 97(8), December 2002, pp. 1213-1220 Production of Amastigotes from Metacyclic Trypomastigotes of Trypanosoma cruzi Víctor T Contreras/+, María C Navarro, Ana R De Lima, Rosa Arteaga, Francy Duran, José Askue*, Yunaimy Franco
Laboratorio de Protozoología,
Centro BioMolP, Facultad de Ciencias de la Salud, Universidad de Carabobo, Bárbula,
Valencia, Estado Carabobo, Venezuela *Facultad de Agronomía, Universidad
Central de Venezuela, Maracay, Estado Aragua, Venezuela This research was supported by grants from Fonacit S1-97000664 (VTC, ARDL), Fonacit S1-2001000683 (MCN, VTC, ARDL), Codecih FCS-97018 (ARDL, VTC) and Codecih FCS-99010 (MCN, VTC). Received 15 May 2002 Code Number: oc02260
Attempts to recreate all the developmental stages of Trypanosoma cruzi in vitro have thus far been met with partial success. It is possible, for instance, to produce trypomastigotes in tissue culture and to obtain metacyclic trypomastigotes in axenic conditions. Even though T. cruzi amastigotes are known to differentiate from trypomastigotes and metacyclic trypomastigotes, it has only been possible to generate amastigotes in vitro from the tissue-culture-derived trypomastigotes. The factors and culture conditions required to trigger the transformation of metacyclic trypomastigotes into amastigotes are as yet undetermined. We show here that pre-incubation of metacyclic trypomastigotes in culture (MEMTAU) medium at 37°C for 48 h is sufficient to commit the parasites to the transformation process. After 72 h of incubation in fresh MEMTAU medium, < 90% of the metacyclic parasites differentiate into forms that are morphologically indistinguishable from normal amastigotes. SDS-PAGE, Western blot and PAABS analyses indicate that the transformation of axenic metacyclic trypomastigotes to amastigotes is associated with protein, glycoprotein and antigenic modifications. These data suggest that (a) T. cruzi amastigotes can be obtained axenically in large amounts from metacyclic trypomastigotes, and (b) the amastigotes thus obtained are morphological, biological and antigenically similar to intracellular amastigotes. Consequently, this experimental system may facilitate a direct, in vitro assessment of the mechanisms that enable T. cruzi metacyclic trypomastigotes to transform into amastigotes in the cells of mammalian hosts.
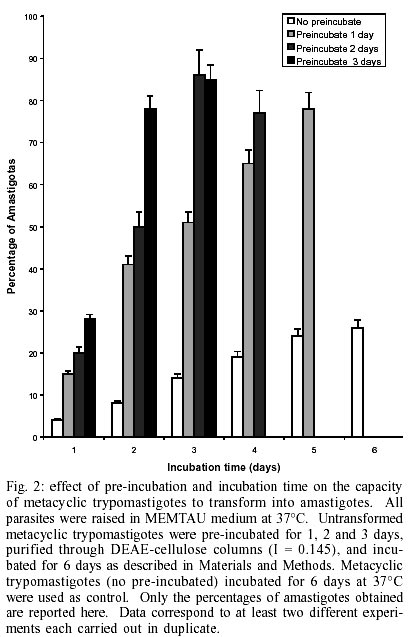
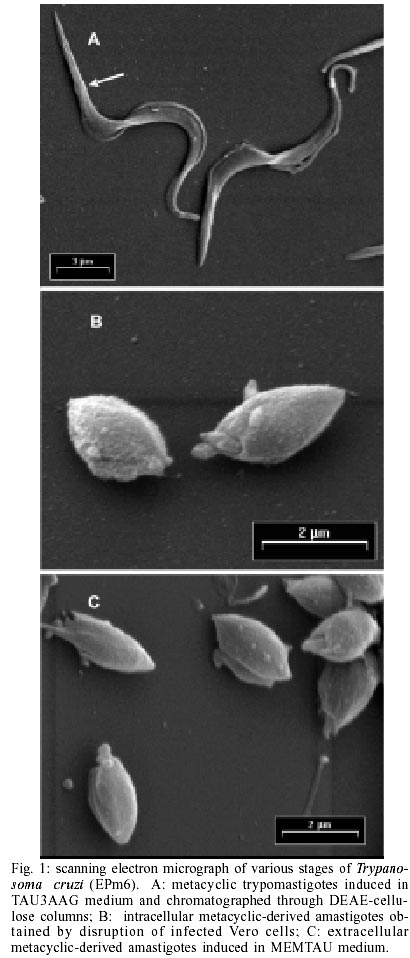
Key words: amastigogenesis - axenic conditions - differentiation - extracelullar amastigotes - morphogenesis Trypanosoma cruzi, the causative agent of Chagas disease, is mainly transmitted to humans by reduviid insects. This parasite has a complex life cycle that alternates between invertebrate and vertebrate hosts. Transmission is initiated by insect vectors, which defecate after a blood meal and release metacyclic trypomastigotes near the bite wound. This infective stage is now capable of reaching an intracellular location and transforming into amastigotes. The amastigotes then divide and transform into trypomastigotes (called bloodstream trypo-mastigotes). Upon leaving the cell, the trypomastigotes enter the bloodstream, where they are eventually taken up by the insect host. In the intestine of the invertebrate host the blood trypomastigote transforms into epimastigotes. The epimastigotes divide and give rise to infective metacyclic trypomastigotes (Brener 1973). Consequently, natural T. cruzi amastigogenesis occurs when metacyclic trypomastigotes from triatomine urine differentiate into amastigotes inside mammalian host cells. Detailed knowledge of the invasion mechanisms of vertebrate cells by T. cruzi has been obtained using cell-culture-derived trypomastigotes that are equivalent to bloodstream trypomastigotes (Burgleigh & Andrews 1995). These studies have revealed that following cell invasion, the trypomastigotes penetrate the host cells. The trypomastigote-to-amastigote transformation is thus initiated, and 1-2 h later, an acid-active membrane pore-forming protein combined with neuraminidase activity synergistically promote the escape of the trypomastigote from the parasitophorous vacuole into the host cell cytoplasm, where the differentiation into amastigote form is completed. Using axenic conditions, several investigators have reported the morphological differentiation of bloodstream trypomastigotes into amastigotes and several factors stimulating this transformation have been described (Villalta & Kierzenbaum 1982, Andrews et al. 1987, Kambara et al. 1990, Tomlinson et al. 1995). Some of these factors are well characterized and represent physiological stress for the parasites. Other authors have obtained extracelullar amastigotes from epimastigotes using highly enriched media supplemented with chicken plasma or vitamins, nucleotides and fetal bovine serum (FBS) (Kimura et al. 1978, Pan 1978, Rondinelli et al. 1988). Although metacyclic trypomastigotes of T. cruzi can be obtained from axenic culture (Chiari 1975, Contreras et al. 1985), it has not been possible to obtain amastigotes in axenic conditions from metacyclic trypomastigotes. Little is known about the factors that control primary amastigogenesis. One obvious difference between a warm-blooded vertebrate and an invertebrate is temperature, and this has been recognized as a morphogenetic element for parasitic protozoa that use arthropods as transmission vectors (Shapira et al. 1988). Since there is difference of approximately 10 degrees between the invertebrate and vertebrate hosts, it is likely that temperature may play an important role in triggering primary amastigogenesis in T. cruzi. For example, even though mammalian cells cultured at room temperature can interiorise metacyclic forms of T. cruzi, transformation of the parasite into amastigotes is not observed (Deane et al. 1984). However, other undetermined factors must exist as most reported experimental designs have failed to recapitulate T. cruzi's primary amastigogenesis (Tyler & Engman 2001). From a parasitological perspective, metacyclic trypomastigotes and bloodstream trypomastigotes appear to share similar biological properties. Both stages are capable of binding to the target cell's membrane, active penetration, transformation into amastigotes, intracellular multiplication and further differentiation into trypomastigotes. However, metacyclic forms and bloodstream trypomastigotes are known to display stage-specific antigens and actually display different modes of interaction with host cells (Burgleigh & Andrews 1995). For instance, metacyclic trypomastigotes express several glycoproteins on their surface that interact with mammalian cells, and which have no counterpart in blood try-pomastigotes (Ruiz et al. 1993). This difference in gene expression justifies the discrimination between bloodstream and metacyclic trypomastigotes as physiologically distinct life cycle stages (Tyler & Engman 2001). In our current work, we describe an experimental protocol for inducing the transformation of T. cruzi metacy-clic trypomastigotes into amastigotes. We have studied the influence of both temperature and pre-incubation times on axenic metacyclics and have found that the incubation of parasites in MEMTAU medium at 37ºC can induce amastigogenesis. Furthermore, we provide morphological, biological and antigenic evidence demonstrating that the in vitro-generated amastigotes are similar to intracellular amastigotes. MATERIALS AND METHODS Parasites and stages - Throughout this study, cloned EPm6 isolate of T. cruzi was used. The parasite was cloned and maintained by alternate triatomine/mouse passages as previously described (Contreras et al. 1994). Epi-mastigotes were cultured at 27°C in LIT medium, and purified metacyclic trypomastigotes (META) were obtained from TAU3AAG medium as previously described (Contreras et al. 1985, Bonaldo et al. 1988). Intracellular derived metacyclic amastigotes (IMA) were purified from monolayer of Vero cells 72 h after infected with metacyclic trypomastigotes. Infected monolayers were gently tripsynised, and the detached cells were recovered by centrifugation and re-suspension in PBS (0.15M sodium chloride, 0.02M sodium phosphate, pH 7.2) supplemented with 1% BSA (bovine serum albumin) (PBS-BSA). A 10 ml aliquot of the re-suspension was disrupted by passage through a 25-gauge needle (Piras et al. 1982). The intracellular amastigotes were separated from the cell debris by centrifugation in a 15-21% discontinuous Me-trizamide gradient (Carvalho & De Souza 1983). Extracellular derived metacyclic amastigotes (EMA) were obtained from metacyclic trypomastigotes incubated in MEMTAU medium as described below. META, IMA and EMA were washed twice with PBS, concentrated by centrifugation in Eppendorff tubes (12,000 xg, 5 min 4°C). Wet masses of parasites (20-25 mg) were stored at -70°C until use. In vitro amastigogenesis - DEAE-52-purified me-tacyclic trypomastigotes (PSG: 0.073M NaCl; 0.005M sodium phosphate; 1% Glucose, pH 8.0; ionic strength (I) 0.181) were concentrated by centrifugation and re-suspended in cold MEM medium without serum (1.0 to 1.5 x 109 parasites/ml). One ml of re-suspended parasites was transferred to sterile plastic tissue culture flasks (175 cm2; Falcon Labware, Oxnar, CA) containing 14 ml of MEMTAU medium pH 5.8, which consists of a 1:1 mixture of TAU3AAG medium and MEM 10% FBS medium, supplemented with 70 mM sucrose, 20 µg/ml bovine or human hemoglobin, 200 U/ml penicillin, 200 µg/ml streptomycin, and 20 mM MES [2 (N-morpholinoethanesulfonic) acid hydrate], followed by incubation at 37°C without agitation in a 5% CO2 atmosphere for three days as the pre-incubation phase. After three days of pre-incubation, the parasites were centrifuged and re-suspended in PSG (I = 0.145), washed once, and applied to the top of a DEAE-52 (diethylaminoethyl) column previously equilibrated with PSG (I = 0.145). Parasites were eluted with 20 ml of PSG (I = 0.145) at room temperature. Aliquots of eluates containing 100% metacyclic trypomastigotes were concentrated by centrifugation (8,000 xg, 10 min, and 4°C) and re-suspended in fresh MEMTAU medium. Parasite suspensions were transferred to sterile culture flasks containing 15 ml of MEMTAU medium (final concentration of ~ 4 x 107 forms/ml), followed by incubation at 37°C (re-incubation phase) for different times. The in vitro differentiation was monitored by counting cells in a hematocytometer and by differential counting in Giemsa-stained smears. The protocols performed in experiments to study the effect of temperature or incubation times are described in the legends of the Table and in Fig. 2. For mass production of EMA (~ 100%), parasites re-incubated for three days were centrifuged as described, and differentiated forms were dispersed in the same volume of fresh MEMTAU medium and incubated at 37°C for 48 h. Scanning electron microscopy (SEM) - Sample processing was carried out essentially as described by Andrews et al. (1987) using glass cover slips precoated with 10 µg/ml poly-L-lysine. Glutaraldehyde-fixed parasites were post-fixed with 1% OsO4 in sodium cacodylate buffer, dehydrated in graded ethanol and critically point dried from liquid CO2 without the use of a transition fluid. Specimens were coated with gold-palladium in a BAL-TEC SCD050 evaporator before being examined in a Philips XL-20 scanning electron microscopy. Lysis of parasites - Wet masses of parasites were suspended (25 mg/100 µl) in lysis solution (150 mM NaCl, 10 mM Tris/HCl pH 7.4, 1 mM EDTA, 1 mM Iodoaceta-mide, 1 mM 1,10-phenanthroline, 0.1 mM PMSF (phenyl-methylsulfonyl fluoride), 0.5 mM TLCK (tosylly-sylchloromethylketone), 0.5 mM TPCK (tosylphenyl-alanylchloromethylketone), 0.1 mM E-64 [L-trans-epoxysuccinylleucyl-amido(4-guanidino)-butane], 25 µM leupeptin, 10 µg/ml antipain, 0.5% NP-40), frozen quickly in dry ice-ethanol, thawed in a 37°C water bath and vigorously vortexed for 3 min at room temperature. After three cycles of freezing and thawing, the disrupted parasites were centrifuged at 12,000 g for 5 min. The supernatants were collected and stored in aliquots, which were either used immediately or frozen for storage at -70°C in 5% glycerol. The protein contents of the supernatants were quantified using the Coomassie Plus assay (Pierce, Rockford, Illinois). Analysis of polypeptides and glycopeptides by sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) - The proteins were analysed by electrophoresis on a linear gradient of 7-15% polyacrylamide gels (Laemmli 1970). Wells were loaded with equivalent amounts of protein: 12 µg for polypeptides, and 20 µg for glycopeptides. The resulting protein patterns were stained with a combined Coomassie Blue-Silver staining procedure (De Moreno et al. 1985) for proteins, and combined Periodic Acid-Alcian Blue-Glutaraldehyde-Silver (PAABGS) stain procedure for glycoproteins. Oxidation with periodic acid and subsequent Alcian Blue prestaining was performed according to Moller and Poulsen protocol (Moller et al. 1993, 1995). Silver staining of glycoproteins and reversal silver staining of proteins in the polyacrylamide gel was achieved by treating the gel with glutaraldehyde. The method of Dubray and Bezard (1982) was used except that 5% aqueous Glutaraldehyde solution was employed instead of 10% Glutaraldehyde buffered. The gel was stained at room temperature for 45 min with 0.1% aqueous silver nitrate rather than 0.4% aqueous silver nitrate at 40°C for 6.5 min. All the steps were carried out with mild agitation. Two types of markers were run in the same gel: the wide molecular weight range used to estimate molecular weight and to control the reversal silver staining (unstaining) of proteins; and a mixture (1 µg/µl) of proteoglycans and glycoproteins (6 µl/well). The following glycoconjugates were used: Chondroitin 6-Sulfate from shark cartilage (type C, Sigma C-4384), Mucin from bovine sub-maxillary glands (type I-S, Sigma M-3895), Fetuin from fetal calf serum (Sigma F-2379), and Bovine a-Acid Glycoprotein (orosomucoid, Sigma G-9014). The gels were then scanned in a Bio-Rad Imaging Densitometer, model GS-690, and their profiles were analysed using the Bio-Rad Molecular Analyst®/PC 1.2 software package. Antisera production and Western blot analysis - Metacyclic trypomastigote stage-specific antiserum (anti-META) was prepared as previously described (Contreras et al. 1998). Antiserum directed against intracellular metacyclic-derived amastigotes (anti-IMA) was prepared in New Zealand rabbits as follows. Twenty mg (wet weight) of parasites was suspended in 2 ml of Freud's complete adjuvant and injected in rabbits in intervals of 8 days, first subcutaneously in the back, and then twice intramuscularly. After two weeks, the rabbits were inoculated intravenously three times at 48 h intervals with 2 mg of freeze-thawed parasites in saline solution. Two weeks after the last inoculation, the immune response was assayed by indirect immunofluorescense with the homologous antigen. Three or four weeks after the last inoculation, the rabbits were bled and the antiserum obtained was kept in aliquots at -70°C. For Western blot analyses, total proteins of META, IMA and EMA were electrophoretically transferred from a linear gradient of 7-15% SDS-PAGE gels to nitrocellulose membranes according to the method of Towbin et al. (1979) as previously described (Contreras et al. 1998). Four µg of protein of each stage were applied to the lanes. The membranes were blocked and incubated overnight at 4°C with a 1: 8,000 dilution of anti-META serum or 1:5,000 dilution of anti-IMA serum. The antigens were revealed using peroxidase-conjugate goat anti-rabbit total immunoglobulins (1:10,000) and the Luminol chemiluminescense system according to the protocol supplied by the manufacturer (Super-Signal, Pierce). Finally, the blots were wrapped in plastic wrap and luminographed using AGFA films. The immunoblots were then scanned and their profiles were analysed as described above. RESULTS In vitro transformation of metacyclic trypomastigotes into amastigotes - At 37°C, MEM 0.4% BSA medium has been reported to induce 100% of bloodstream try-pomastigotes into amastigotes (Tomlinson et al. 1995). However, under these conditions metacyclic try-pomastigotes failed to produce amastigotes, and after one day of incubation all the parasites lysed. After testing different media and incubation conditions, we could obtain a high yield of differentiation forms and EMA at 37°C in a 5% CO2 atmosphere. This was achieved in two steps: Step 1 consisted of the pre-incubation of metacyclic trypomastigotes at 37°C in MEMTAU for three days. The metacyclic trypomastigotes underwent very few morphological changes, and a high percentage of well-preserved metacyclic trypomastigotes (up 80%) was obtained; Step 2: the subpopulation of untransformed metacyclic trypomastigotes was purified through DEAE-52 columns (I = 0.145), and when these purified metacyclic try-pomastigotes were incubated again in fresh MEMTAU medium at 37°C, a high percentage (up 80%) of typical, well-preserved amastigotes were progressively produced. The in vitro-derived amastigotes did not display any detectable signs of degeneration and were indistinguishable from normal amastigotes. In vitro generated amastigotes are morphologically indistinguishable from naturally produced amastigotes - Fig. 1 shows the morphology of metacyclic forms and compares EMA and IMA by SEM. Metacyclic trypomastigotes that had recently been transformed from epimastigotes and purified through a DEAE-52 column (I = 0.181) showed a slender morphology and displayed a subterminal kinetoplast (arrow, Fig. 1A). General aspects of the EMA and IMA are shown in Fig. 1B, C, respectively. The comparison of EMA and IMA isolated by Metrizamide gradient showed a remarkable morphological similarity. They were rounded or oval-shaped and a short protruding flagellum was observed. Further characterization of the transformation process will be given elsewhere (Contreras et al., in preparation). Effect of temperature, pre-incubation and incubation and age of the culture media on the in vitro production of amastigotes from metacyclic trypomastigotes - We wanted to test which other factors affected the in vitro transformation process. When metacyclic trypo-mastigotes were pre-incubated at 37°C, purified by DEAE-52 columns, and incubated in fresh medium at 37°C a high percentage of amastigotes (85 ± 6%) was observed (Table). However, when they were pre-incubated at a lower temperature (27°C), purified and placed at 37ºC, only 25 ± 5% of amastigotes was observed. In addition, incubation of metacyclic trypomastigotes in fresh medium at 27°C, followed by purification and incubated at the same temperature (27°C) did not affect parasite morphology (100% metacyclic trypomastigotes). On the other hand, when temperature-committed metacyclic trypomastigotes were incubated in fresh medium at 27°C for 3 days, the pattern of morphologic changes showed a higher percentage of differentiation forms (38 ± 7%) than amastigotes (10 ± 4%), indicating that this metacyclic trypomastigote population could not complete the morphological transformation at the lower temperature. Purification of parasites through DEAE-52 had no effect on committing metacyclic trypomastigotes to amastigogenesis. Parasites that were pre-incubated for three days at 37°C and then transferred to fresh media without passing through DEAE-52 columns were still capable of producing large numbers of amastigotes (80 ± 3%, Table ). It is therefore unlikely that the purification step modulates the parasite's ability to undergo amastigogenesis in any significant way. Finally, the variables of parasite titer, volume and quality of the incubation medium were tested. Modifications in the inoculation titer ranging from 5 x 106 to 7 x 107 parasites/ml; pH (5.3 to 6.9) and the depth of medium (1 to 7 mm) had no significant effect on the transformation process (data not shown). However, we noticed that amas-tigotes production is markedly compromised in old medium. For instance, if the same pre-incubation medium used to "pre-condition" the metacyclic trypomastigotes is used again for incubation, a fairly low number of amastigotes were obtained (57 ± 6%, Table ) in comparison with parasites incubated in fresh medium. In all likelihood, this may be due to the progressive exhaustion of nutrients and/or catabolite accumulation in the media. During the course of experimentation, it became evident that temperature (37ºC) is crucial to allowing metacyclic trypomastigotes to become competent and complete the amastigogenesis process. In parasite-vertebrate cell culture systems, it is known that following penetration, metacyclic trypomastigotes differentiate into amastigotes in approximately 3 h and remain morphologically quiescent for about 35 h (at 35°C) prior to the onset of reproduction (Dvorak 1975). If the observed temperature-induced transformation reported here has physiological significance, pre-incubation at 37°C for different times should affect the number of metacyclic trypomastigotes committing to amastigogenesis. To test this hypothesis, metacyclic trypomastigotes were pre-incubated at 37°C for 1, 2 and 3 days. The parasites were later purified and incubated in fresh MEMTAU medium. Their respective transformation kinetics were measured during the following six days of culture. Metacyclic trypomastigotes incubated for six days at 37°C without pre-incubation were used as control. Fig. 2 shows the effect of pre-incubation length on the capacity of metacyclic trypomastigotes to transform into amastigotes. The transformation process was accelerated when the metacyclic trypomastigotes were pre-incubated for 1, 2 or 3 days. Parasites pre-incubated for a single day yielded 51 ± 1% and 78 ± 5% of amastigotes after three or five days of incubation, respectively. When pre-incubated for two days, amastigotes were generated at a yield of 86 ± 4% and 77 ± 6% at the end of days three and four of incubation. Finally, if the parasites were pre-incubated for 3 days, the percentage of amastigotes rapidly increased to 85 ± 6% toward the end of the third day of incubation. It is important to note that the control condition (not pre-incubation) showed a steady increase in the percentage of amastigotes produced (14 ± 3% and 26 ± 3% during days 3 and 6, respectively), but never reached the numbers obtained in the pre-incubation experiments. These results indicate that pre-incubation at 37°C induces a commitment of metacyclic trypomastigotes to differentiation and indicates that under our experimental conditions (without steady-state) pre-incubation for two days was sufficient to produce large number of amastigotes. Thus, the data on culture conditions suggest that in vitro amastigogenesis occurs in two phases: a temperature-dependent induction of transformation, and an initiation of morphological transformation which is dependent on temperature and the availability of fresh, protein-rich medium. In vitro generated amastigotes
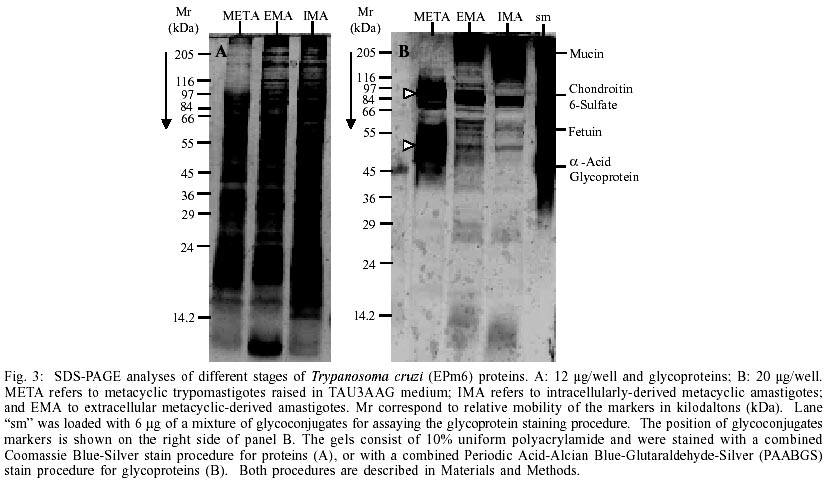
show a high degree of similarity of polypeptides and glycopeptides with the
naturally produced amastigotes - With the purpose of making a molecular-level
comparison of the stages produced, total proteins from META, EMA and IMA were
analyzed by SDS-PAGE, and their polypeptide profiles were visualized by Coomassie
Blue-Silver as shown in Fig. 3A. Two distinct
protein profiles can be observed. One displayed by TAU3AAG-purified metacyclic
trypomastigotes before incubation in MEMTAU medium at 37°C (lane META,
Fig. 3A) and another shared by both amastigotes
(EMA and IMA). It is worth noting that EMA displayed a protein profile significantly
different to that of META, and that the profiles of IMA and EMA show surprising
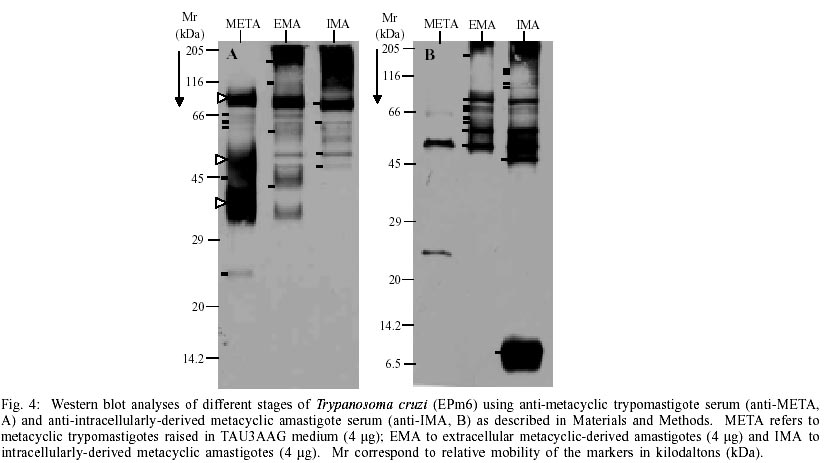
similarities between the 205 and 14.2 kDa To determine whether there were changes at the glycoprotein level, as observed in the protein profiles, an equivalent gel was visualised by the PAABGS procedure as shown in Fig. 3B. The patterns of glycopeptides obtained are characteristic for each morphological stage. Metacyclic trypomastigotes showed a pattern characterized by a major glycopeptide band with a molecular mass ranking from 80 to 92 kDa, and the presence of a smear of glycopeptides with a molecular mass ranking from 47 to 57 kDa (white triangles at left of lane META, Fig. 3B). The comparison of this profile with the glycoprotein profile displayed by EMA shows significant changes, represented by the appearance of an intensely silver-stained cluster of high molecular weight, unresolved glycopeptides migrating as the Mucin marker, which is absent in META, a rearrangement of glycopeptides between the 66 and 97 kDa markers with a major glycopeptide band of molecular mass ranking from 75 to 85 kDa and the appearance of at least 5 well-resolved glycopeptides of similar intensity between the 66 and 45 kDa markers. A comparison of EMA and IMA patterns shows remarkable similarities. These results indicate that the morphological transformation of the parasites is associated with specific changes in their polypeptides and glycopeptides profiles and may reflect the degree of similarity of protein content shared by both types of amastigotes. Western blot analysis of stage-shared antigens - Stage-specific antigens were used to determine if the changes observed in META, EMA and IMA proteins and glycoprotein profiles reflected specific morphological changes. Western blots of all three stages were performed using stage-specific antisera. Fig. 4A shows antigen profiles of META, EMA and IMA revealed by anti-META serum. Metacyclic trypomastigotes showed a pattern characterized by two dark clusters of unresolved antigens, a smear (95/80, 43/33 and 52/46 kDa, respectively, white triangles at left of lane META, Fig. 4A) and five barely detectable antigens (66, 64, 60, 45 and 25 kDa, short bars at left of lane META, Fig. 4A). The comparison of this profile and the one displayed by EMA shows significant changes represented by the appearance of at least four antigenic bands (> 175 cluster, 115, 58 and 43 kDa, short bars at the left of lane EMA, Fig. 4A) which are absent in META. Four antigenic bands were recognized by anti-META serum in META, EMA and IMA (82, 64,52 and 47 kDa, short bars at the left of lane IMA, Fig. 4A). The comparison of EMA and IMA patterns shows similarities (Fig. 4A). Finally, the antigenic profiles of META, EMA and IMA revealed by anti-META serum (Fig. 4A) resemble the glycopeptide profiles of these stages (Fig. 3B), suggesting that almost all of the glycosilated molecules of the metacyclic trypomastigotes are highly immunogenic. Stage comparisons using anti-IMA serum confirmed the remarkable similarity between both amastigotes studied (EMA and IMA, Fig. 4B), which share at least eight antigenic bands of different width and density (> 175, 75, 72, 70, 64, 60, 55 and 51 kDa, short bars at the left of lane EMA, Fig. 4B). Likewise, META was poorly recognized by anti-IMA (Fig. 4B). It should be noted that the 47 kDa and cluster 12/5 kDa and several antigenic bands (135, 120, 105 and 95 kDa) in the region of the 116 kDa marker revealed by anti-IMA in IMA (short bars at left of lane IMA, Fig. 4B) were not present in the other two stages, suggesting that such antigens may be IMA-specific. Therefore, the immunoblotting data indicate that the morphological transformation of the parasites are most likely associated with antigenic changes and are likely to reflect stage-specific molecular profiles. Similarly, these data also suggest that common antigens are shared by metacyclic and amastigotes stages. DISCUSSION
One of the main findings of the present study is that amastigogenesis can be triggered in vitro by temperature changes in media supplemented with proteins (serum). The practical extension of this result being the development of a system that allows the in vitro production of high yields of extracellular T. cruzi amastigotes from metacyclic trypomastigotes. Most of the information about factors triggering in vitro amastigogenesis has been obtained by assaying bloodstream trypomastigotes in which the parasites are neither subjected to a heat shock nor to density changes in their environment. It is known, for instance, that low pH accelerates the extracellular transformation of tissue culture-derived trypomastigotes into amastigotes (Kambara et al. 1990, Tomlinson et al. 1995). However, metacyclic trypomastigotes incubated under identical conditions do not differentiate but rather undergoes cell death over a 24-hour period (Tyler & Engman 2001). This fact seems to indicate that metacyclic trypomastigotes posses a specific way of responding and adapting to environmental changes that is different to that shown by bloodstream trypomastigotes and may explain the previous inability to produce amastigotes axenically. Several authors have devised cell-free media in which a mixture of epimastigotes and metacyclic trypomastigotes from cultures of T. cruzi are induced to transform into amastigotes (Pan 1978, Kimura et al. 1978, Villalta & Kierzenbaum 1982, Rondinelli et al. 1988). In such assays, the amastigotes were gradually selected by employing highly enriched media and a progressive increase of the incubation temperature by serial culture at 35.5°C or 37°C. The approach described in this report does away with the use of developmentally heterogeneous populations of parasites to produce amastigotes. Instead, it relies on a homogeneous, purified metacyclic trypomastigote population raised in a chemically-defined medium at 27°C. Pre-incubation of this axenic metacyclic population for at least two days at 37°C in a protein-supplemented medium becomes amastigogenic when cultured again in fresh, protein-rich medium (Fig. 2). Furthermore, the addition of a DEAE-cellulose purification step to remove differentiation forms and enrich metacyclics after pre-incubation did not affect their ability to undergo amastigogenesis. We can not rule out the possibility that specific proteins present in sera might act as triggers for the differentiation process. However, parasites pre-incubated in media (without serum) and supplemented with hydrolyzed lacto albumin and incubated in MEMTAU medium also underwent the reported transformation process, although to a lesser extent (data not shown). We therefore believe that the "pre-conditioning" serves to exert physical changes on the purified metacyclic trypomastigote, resulting in measurable intrinsic changes in the biological properties of the parasite. Future experiments will test whether culturing the parasites in media of different specific densities is sufficient to trigger amastigogenesis on its own. We also demonstrate that the EMA produced in vitro are similar to IMA using several different criteria (gross morphology, protein and glycoprotein profiles as well as immunogenic profiles). In fact, preliminary data show that the in vitro produced EMAs are not only capable of infecting cultured cells and mice, but are also resistant to macrophage digestion (data not shown). Recently, we have studied the changes that occur at molecular level during the in vitro metacyclic trypomastigote-amastigote transformation. We founded that the in vitro primary amastigogenesis is associated with abrupt changes in protein, glycoprotein, and stage-specific antigens that occur simultaneously during the first 24 h of pre-incubation (Contreras et al. 2002). Therefore, based on the results reported here, we postulate that at least two factors are involved in triggering primary amastigogenesis: the increase in temperature and the changes in viscosity encountered by metacyclic trypomastigotes upon invasion of the vertebrate host. It has been reported that the heat-shock proteins (hsp) could play an important role in the infection and differentiation of T. cruzi (Requena et al. 1992). However, the effect that the rheologycal properties of the transmission fluids may play in the T. cruzi life-cycle have yet to be systematically studied and should be explored more extensively. In conclusion, this paper reports for the first time the in vitro production of extracellular amastigotes from axenically-reared metacyclic trypomastigotes. It is still premature to assume that our in vitro differentiation system precisely recapitulates naturally occurring amastigogenesis, i.e., the transformation of metacyclic trypomastigotes from triatomine urine into amastigotes inside the cells of mammalian hosts. However, preliminary data indicate that in vitro produced amastigotes cannot be distinguished from intracellular, metacyclic-derived amastigotes obtained from Vero cells in terms of their biological properties (Contreras et al., in preparation). One clear advantage of our experimental system is that it makes use of in vitro produced metacyclic trypomastigotes, which can be massively obtained from chemically defined media. Thus, EMA obtained in vitro could serve to delineate the molecular differences and similarities that must exist between amastigotes and trypomastigote-derived amastigotes, effectively providing a simple and inexpensive alternative for the identification and testing of new chemotherapeutic and immune prophylactic agents.
ACKNOWLEDGEMENTS
To Dr Alejandro Sánchez for his critical suggestions and helpful discussion of the manuscript. To Gregorio Flores and Mr Wilmer Pineda for their excellent technical assistance. REFERENCES
Copyright 2002 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc02260f3.jpg] [oc02260f2.jpg] [oc02260f4.jpg] [oc02260f1.jpg] [oc02260t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}