 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98(Suppl. I), January 2003, pp. 29-34 The Fossil Tabanids (Diptera Tabanidae): When They Began to Appreciate Warm Blood and When They Began Transmit Diseases? Rafael Gioia Martins-Neto
Sociedade Brasileira de Paleoartropodologia, Universidade Guarulhos, Rua Arnaldo Vitaliano 150, apto 81, 14091-220 Ribeirão Preto, SP, Brasil, Fax: +55-16-624.2884. E-mail: martinsneto@terra.com.br Received 26 August
2002 Code Number: oc03006 A discussion of the known fossil tabanids (Diptera Tabanidae) is presented based on fossil evidence. This includes the origin of the hemathophagy in the Brachycera, more specifically for tabanids. Several tabanid species in the extant fauna are vectors for disease-producing organisms that affect humans and animals. Bacteria, viruses, rickettsiae, protozoa, and filarial worms can be transmitted by them, causing such diseases as anthrax, tularemia, anaplasmosis, various forms of trypanosomiasis, Q fever, and filariasis. However, if tabanids are directly responsible for all of these diseases is not consensual and the known fossil evidence is presented here.
Key words: Diptera - fossil Tabanidae - paleoparasitology - blood-suckers evolution - diseases in the past The bloodsucking insects of the orthorraphous families of Diptera are known from the Early Cretaceous and include Psychodida, Corethrellidae, Culicidae (with some doubts), Ceratopogonidae, Simuliidae, Rhagionidae, Tabanidae, and Athericidae (Ansorge 1994, Lukashevich & Mostovski et al. 2001). Although brachycerous flies are abundant in the fossil record, with several families recorded (Apystomyiidae, Archocyrtidae, Asilidae, Eremochaetidae, Hilarimorphidae, Kovalevisargidae, Mythicomyiidae, Nemestridae, Protapioceridae, Raghionempididae, Empididae, Scenopinidae, Stratyomyidae, Therevidae, Vermileonidae, Xylomyidae, and Xylophagidae), since the Middle Jurassic (Mostovski 1998), tabanids are relatively scarce in the fossil record (mainly in Mesozoic). The older record of a true tabanid came from the Upper Jurassic deposits of China (Ren 1998), with three genera and three species, all related to the flower feeders, Pangoniinae. The Cretaceous record comprises specimens from the Lower Cretaceous of England (Coram et al. 1995) and Spain (Lukashevich & Mostovski 2001), and a possible specimen in the Cretaceous of South Africa (Mostovski 1998). For the Cenozoic, the following named species are known: from the Miocene of Florissant, Colorado, respectively named Tabanus parahippi Cockerell 1909, Tabanus hipparionis Cockerell 1909, and Tabanus merychippi Cockerell 1917; from the North American Oligocene, Silvius merychippi Melander 1947; from the Germany Oligocene, Tabanus statzi Moucha 1972; from the French Olicocene, Aemodipsus bornensis Maneval 1936 and Chrysops seguyi Piton 1940; from the Swittzerland Oligocene, Hexatoma oeningensis (Herr 1864) Evenhuis 1994, and Tabanus vectensis Cockerell 1921, for the England Eocene/Oligocene. From the Baltic amber (Eocene/Oligocene), Stenotabanus brodzinskyi Lane, Poinar & Fairchild 1988 and Stenotabanus woodruffi Fairchild & Lane 1989. Tabanidae were also found in Oligocene inclusions from Chiapas, Mexico (Morales & Pimentel 2001). For the Pliocene from Poland, Europe, North Africa and Morocco the species Tabanus sudeticus Zeller 1842 was recorded. An unnamed species was assigned to the genus by Grabenhorst 1985, for the German Pliocene. Scudder (1895) recorded a possible new genus and species of tabanid from the Miocene Oeningen deposits of Switzerland. Undescribed Tabanidae have been noted from the Eocene of the Green River Formation (Bradley 1931, Swanson & Lewis 1993). The Cretaceous genus Protabanus, described by Hong (1982) has been shown by Grimaldi (1990) and Zhang (1993) to belong to the order Homoptera, where it has been placed in the cicadoid family Tettigarctidae by Hamilton (1992) in his catalog of Mesozoic Homoptera genera (see also Evenhuis 1994). The species of Haematopota described by Stuckenberg (Haematopota pinicola Stuckenberg 1975) as from Baltic amber has been shown by Lewis et al. (1977) to actually be a copal inclusion from an unknown locality in Africa. An older undetermined compression fossil of Tabanidae from the Middle Jurassic Mont-Saint-Marin deposit in Luxemburg was noted by Maurice (1882). There are Cretaceous genera described by Kovalev (1986) from Mongolia, but are treated by Evenhuis (1994) as unplaced in Tabanoidea. THE BRAZILIAN FOSSIL DIPTEROFAUNA
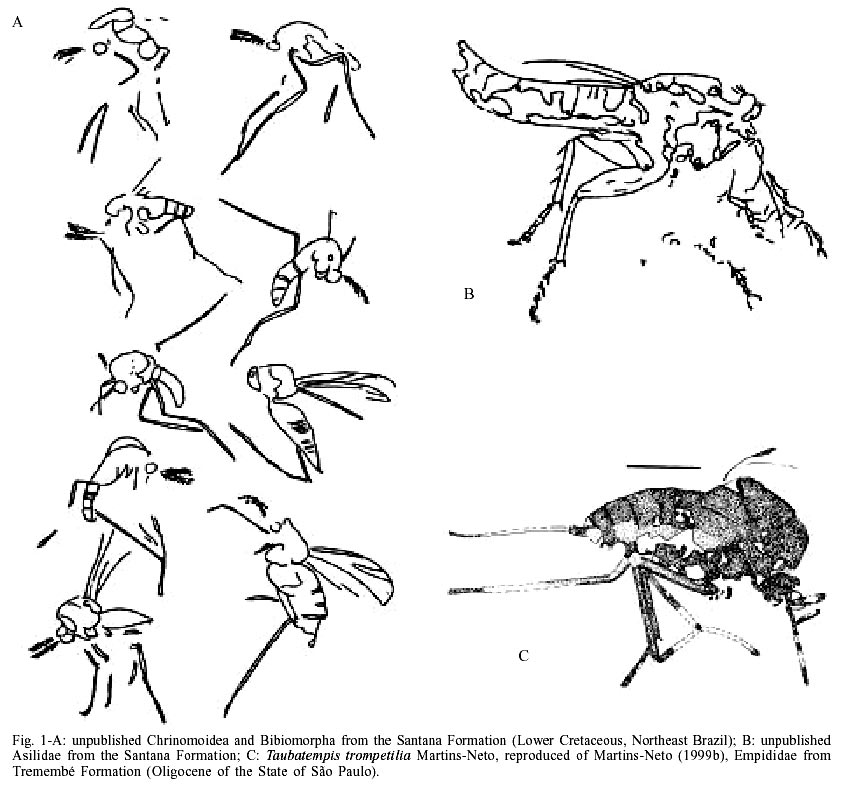
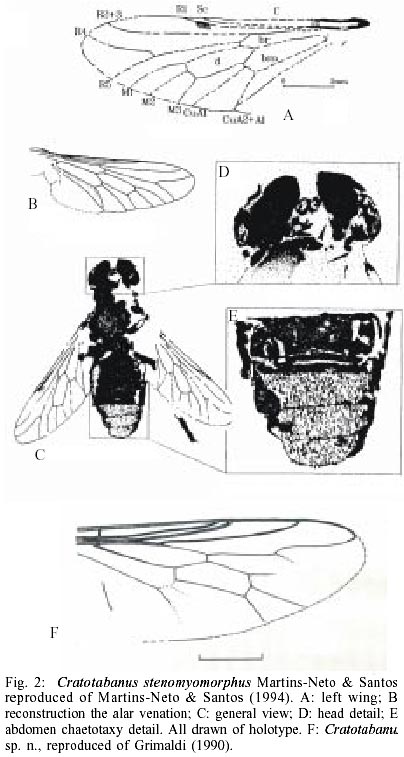
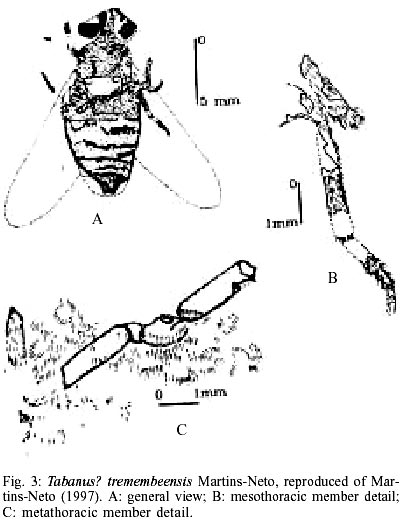
The presently known Brazilian dipterofauna came just from three localities: Santana Formation (Lower Cretaceous, Northeast Brazil), Tremembé Formation and Entre-Córregos Formation (both Oligocene, Southeast Brazil). From the Santana Formation the following Diptera groups are known: Chironomoidea (unpublished material, Fig. 1A), Asilidae, (Grimaldi 1990, and further unpublished material - Fig. 1B), Bibiomorpha (unpublished material -Fig. 1A), Tipulidae (Ribeiro & Martins-Neto 1999, Ribeiro & Krzeminski 2000), Simuliidae (Vulcano 1985). For the purposes of this contribution it is irrelevant if the Vulcano's brief communication has serious problems with the International Zoological Nomenclatural Code. Of course, it is another nomina nuda in the paleontological literature, because this specimen never receive a systematic treatment. Fortunately I saw this specimen myself and I can confirm that it is a true female simuliid, the oldest known record of a Diptera Simuliidae), and Tabanidae, Cratotabanus stonemyomorphus Martins-Neto & Santos 1994 (Fig. 2). From the Tremembé Formation, Taubaté Basin, Oligocene of the State of São Paulo, are known the following groups: Hybotidae, Mycetophilidae, Diastatidae, Empididae (Fig. 1C), Tipulidae and Tabanidae (Martins-Neto 1997, 1999a, b, Martins-Neto et al. 1992), with the named species Tabanus? tremembeensis Martins-Neto1997 (Fig. 3). From the Entre-Córregos Formation, Aiuruoca, Oli-gocene of the State of Minas Gerais, just a Mycetophilidae species is known. Grimaldi (1990) illustrates a dipteran specimen, interpreted as a Therevidae indet. (Fig. 2D), from the Santana Formation, but this specimen needs be moved to the genus Cratotabanus Martins-Neto & Santos (1994), constituting a new species of Tabanidae for these deposits (the second one). THE BLOODSUCKING TABANIDS. WHEN DID THEY BEGIN?
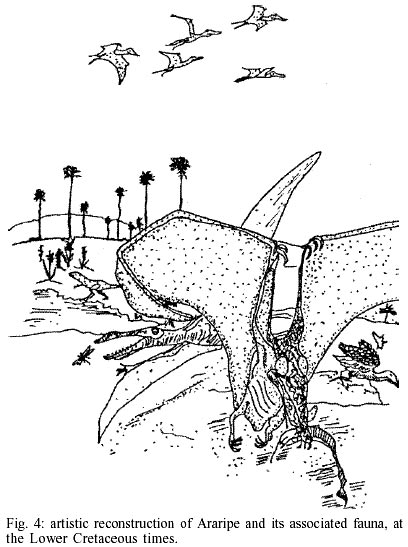
Most adult female Tabanidae, in the extant fauna, feed on blood of warm-blooded animals, but a few species are known to attack cold-blooded vertebrates (Pechuman & Teskey 1981), and several species are not hematho-phagous. These, as well as the females that are blood feeders, and males that are not hematophagous, are known to visit flowers (Pechuman & Teskey 1981), although the emphatic affirmation that tabanid females are, necessarily, hematophagous (Pessôa & Martins 1978), which could sound as an exclusivity for tropical species. However, Martins (1940) had observed Neotropical female tabanids visiting flowers in Minas Gerais, Southeast Brazil (in Verbenaceae, and also in "crisântemos" and orchids), but never he had saw males visiting flowers (just resting in the leaves). Also interesting from the Martins's observations (1940) is the female (hemathophagous) ferociously feed on pigs, often on horses, but rarely on cows, (apart from humans). Chrysops have preference (after the Martin's observations) for the ear region of the mentioned animals. Chrysops rarely feeds on reptiles, such as the Brazilian crocodilid Jacaretinga (common in the State of Mato Grosso). Other Chrysops species feed on the back of horses, others on their legs, and others on their abdomen. Despite the necessity of a rigorous taxonomical review, in the light of the modern systematic methods, the contribution of Martins (1940) revealed the presence of 9 genera and 42 species of Pangoniinae, and 12 genera and 52 species of Tabaninae (the most common genera, Chrysops, with 15 species and Tabanus, with 18 species), just in the Minas Gerais. Martins's main contributions are the pioneer observations on the biology and ecology of neotropical tabanids. Going to the past, the source of warm blood from Santana Formation, cradle of Cratotabanus Martins-Neto, was scarce or insignificant. No mammals were found, at least until now, and just a single avian remain (a feather, Martins-Neto & Kellner 1988) was found, indicating, at least, that the group was represented in the Araripe fauna. There was almost nothing for warm bloodfeeders at this time (at least in a stricto sensu, considering just mammals and aves as true warm-blooded animals). The terrestrial vertebrates of the Araripe fauna were composed mainly of pterosaurs (one of the most important of the world, by the number of species represented as well as by the excellence of preservation), a few dinosaurs, turtles, a few lizards, crocodilians, and birds (an artistic view of the Araripe times is furnished in the Fig. 4). The climate was essentially tropical, optimal for tabanids and several groups of bloodsuckers, with transparent and oxygenated water available which was seasonally toxic with gas (Martins-Neto 1996). Berner et al. (1983) concludes that the CO2 in the atmosphere during the Cretaceous times was greater than the presently. Similar conclusions were obtained by Berner and Landis (1988) through gas bubble inclusions in amber (Cretaceous from Cedar Lake, USA). On this theme, Emiliani et al. (1981) presents an interesting review of the maximal tolerances of temperature by the animals. The theme furnishes material for discussion. For example, it is debated whether or not the dinosaurs and pterosaurs were warm-blooded. Decades of discussion (see Baker 1986) proved, at least, that specific groups of dinosaurs could be warm-blooded and this could open the way earlier for the Cretaceous warm bloodfeeders. Ponomarenko (1976) had found a flea-like insect preserved adhered in a pterosaur alar membrane, from the Siberian Jurassic. This occurrence absolutely does not prove that the insect was drinking warm-blood, but it is a good paleontological evidence for the beginning of the bloodsucking (cold or warm). For tabanids, the older evidence came from the Chinese Jurassic, with the pangoniine tabanid Palaepangonius eupterus Ren 1998, associated with flower-like structures including a complete body and an associated well-developed long proboscis, providing direct evidence for the mid-Mesozoic diversification within Tabanidae of subclades with nectaring mouthparts, as pointed by Ren (1998) and Labandeira (1998). This important find, however, doesn't furnish evidence for a possible tabanid bloodsucker at this time. No Tabaninae was found in Jurassic times, until now. The panorama for the Tertiary is less debatable. There is no resistance to the idea that the tabanids were drinking warm blood from the ears of ungulates, which lived in herds, and several others groups of mammals. However, no direct paleontological evidence sustained this, until now. On the other hand, the record of tabanids known from several Eocene/Oligocene deposits in the world shows that it is highly probable that the group could have the same feeding habit as the present day tabaninids (including the fossil record of several genera living today). THE DISEASES TRANSMISSION BY TABANIDS: WHEN THEY BEGAN? ARE THE TABANIDS REALLY GUILTY?
Several tabanid species in the extant fauna are vectors for disease-producing organisms that affect man and animals. Bacteria, viruses, rickettsiae, Protozoa, and filarial worms can be transmitted by them, causing such diseases as anthrax, tularemia, anaplasmosis, various forms of trypanosomiasis, Q fever, and filariasis (Pechuman & Teskey 1981). However, if tabanids are directly guilty for all these diseases, it is not consensual. Tularemia was discovered in the United States, transmitted, as initially thought, by the tabanid Chysops discalis Williston. Today we know that the true culprit is an acarine, and this disease is not reported or is unknown for Brazil (Martins 1940), although several Neotropical tabanids have been found infected by similar acarines. Rickettisiae, for example, was recently addressed (Gray 1998) but again, no proof of responsibility for tabanid-associated diseases was directly demonstrated. Several diseases are caused by nematodes. Blaxter et al. (1998) present a phylogenetical approach of the phylum Nematoda. They include considerations of trophical ecology such as: bacteriovore, algivore-omnivore-predator, fungivore, plant parasite, entomopathogen, invertebrate parasite, and vertebrate parasite. They concluded that animal parasitism arose independently at least four times, and plant parasitism three times. Strongylida is an exclusively vertebrate parasite group, as well as Ascaridina, Spirurida, Oxyurida, Rhabditidoidea, Strongyloididade and Trichocephalida. Exclusively entomopathogen are the groups heterorhabditids and Steinemematidae. Exclusively invertebrate parasites are the Rhigonematida and Mermithida. There is controversy about the phylogenetic tree and the position of the clades, and the relationships of the main clades and characters used (Voronov et al. 1998). But, for the scope of this paper, of all 55 branches of Blaxter et al. (1998), just two - Rhigonematida and Mermithida - are exclusively invertebrate parasites. The former was represented in the fossil record, parasiting insects, including tabanids, as briefly documented below. Voigt (1957) described an Eocene larval nematode encysted in a muscle tissue of a beetle. Larsson (1978) discussed parasitic nematodes within chironomids from the Baltic amber mermithids associated with dipterans, including an adult limoniid fly and a female mosquito. Taylor (1935) earlier summarized our knowledge of fossil nematodes, including a mermithid parasite within a German Baltic amber Oligocene chironomid (see also Boucot 1990). Statz (1944) presented mosquitoes and potential disease vectors (Culicidae in mammals) from de Germany Oligocene. Cockerell (1908) and Brues (1923) report Glossina (Muscidae) in beds of Oligocene age (fossils nematode trails was found earliear, in Eocene sediments - see Moussa 1970). This evidence suggests the possibility that trypanosome-ungulate, and other mammal relations may have been in existence for some time back into the past (Boutcot 1990). Dogiel (1964) and Grunin (1973) present Late Pleistocene warble-fly larvae (Cobboldia) occurring in the guts of mammoths from Siberia. Elephants today are also parasitized by these same flies. Specifically for tabanids, the geological record does not furnish direct evidence until the Late Pliocene, when Grabenhorst reports mermithid nematodes infecting a fossil tabanid. The simple presence of fossil tabanids, and possible victims of their bite, falls in Boucot's (1990) Category 6: highly speculative, but the presence of nematodes in tabanids falls in the Boucot's (1990) Category 1: incontestable, the first and single for fossil tabanids. FINAL COMMENTS
The fossil evidence furnishes us the following facts, conclusions, and speculations: (1) Flower-feeding tabanids (Pangoniinae) appear, at least, in the Upper Jurassic. This represents, in a phylogenetical sense, that bloodsucking (for tabanids) is a sinapomorphic condition, maybe attained later than the Lower Cretaceous (based on negative evidence rather than direct evidence); (2) Tabaninae tabanids appear, at least, in the Lower Cretaceous (based in negative evidence), earlier than the Aptian; (3) The presence of fossil tabanines in the Lower Cretaceous of Asia, South Africa, Brazil, Spain and England, in the light of the paleobiogeography, indicates that the group was represented earlier, at least in the beginning of the Cretaceous; (4) In a phylogenetical sense, cold bloodsucking precedes warm bloodsucking, indicating the Lower Cretaceous as the beginning for the bloodsuckers; (5) Warm bloodsucking in tabanids could have started in tropical regions, later than the mid-Cenozoic (speculative, based on negative evidence); (6) Disease transmission by tabanids is highly controversial. Presently, of all diseases attributable to tabanids, just nematoid infection is incontestable. The first paleontological evidence, however, dates from Upper Pliocene. For other dipteran groups the record is older, at least since the Eocene; (7) Tabanids are not guilty of all crimes (diseases) attributable to them. For several accusations there exist reasonable doubts.
ACKNOWLEDGEMENTS
To Dr Reginaldo Peçanha Brazil (Centro de Pesquisas René Rachou-Fiocruz), by the incentive, friendship, criticisms and suggestions; to Dr Deborah Rita Brunherotto (Sociedade Brasileira de Paleoartropodologia) for the inestimable help in pathology, physiology and by the fertile discussion on the theme; to Dr Conrad Chrystopher Labandeira (Smithsonian Institu-tion, USA), a plant-arthropods interactions specialist, by sug-gestions, comments, and criticisms; to Dr Rob Coram (Maidstone Museum, UK) and Dr Ren Dong (National Geo-logical Museum of China), by the friendship and help with reprints. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03006f1.jpg] [oc03006f4.jpg] [oc03006f3.jpg] [oc03006f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}